സൈറ്റിൻ്റെ വിഭാഗങ്ങൾ
എഡിറ്ററുടെ തിരഞ്ഞെടുപ്പ്:
- ഐടി ഉദ്യോഗസ്ഥർ എഞ്ചിനീയറിംഗ്, ടെക്നിക്കൽ തൊഴിലാളികളാണ് (ട്രാൻസ്ക്രിപ്റ്റ്)
- ശിക്ഷിക്കപ്പെട്ട ഒരു വ്യക്തിയുടെ സാമൂഹികവൽക്കരണത്തിൻ്റെ 14 പ്രധാന ദിശകൾ
- വ്യാസെസ്ലാവ് നാഗോവിറ്റ്സിൻ ഒരു ജോലിയും കണ്ടെത്തിയില്ല
- അലക്സി പുഷ്കോവ്: ജീവചരിത്രം, വ്യക്തിഗത ജീവിതം, കുടുംബം, ഫോട്ടോ ബാല്യം, കുടുംബം
- പെച്ചെർസ്കിലെ വിശുദ്ധ സിസോയ്, സ്കീമാമോങ്ക്, പെചെർസ്കിലെ വിശുദ്ധ വെനറബിൾ സ്കീമാമോങ്ക് ജോസാഫ്
- യേശുക്രിസ്തുവിൻ്റെ പുനരുത്ഥാനം ക്രിസ്തുവിൻ്റെ പുനരുത്ഥാനത്തിനുശേഷം സംഭവിച്ചത്
- കത്തീഡ്രൽ ഓഫ് ബ്ലെസ്ഡ് വിർജിൻ മേരി കത്തീഡ്രൽ
- ഓർത്തഡോക്സ് എൻസൈക്ലോപീഡിയ ട്രീയിലെ നാലാമത്തെ മെനയോണിൻ്റെ അർത്ഥം
- ആരോഗ്യ മന്ത്രാലയത്തിൻ്റെ ഉത്തരവ് 290n തീയതി 02
- എക്സ്പോണൻഷ്യൽ അസമത്വങ്ങൾ
പരസ്യം ചെയ്യൽ
| I2 ഹാപ്ലോഗ് ഗ്രൂപ്പ്. സ്ലാവുകളും സബ്സ്ട്രേറ്റും (ഹാപ്ലോഗ് ഗ്രൂപ്പ് I2a1 സംബന്ധിച്ച പരിഷ്കരിച്ച പതിപ്പിൻ്റെ ശകലം) |
|
111-മാർക്കർ ഹാപ്ലോടൈപ്പുകൾ ഏറ്റവും വിശദമായി എടുത്തിട്ടുണ്ട്, അവയിൽ ആദ്യത്തെ 12 മാർക്കറുകളുള്ള 15 ഹാപ്ലോടൈപ്പുകൾ അവതരിപ്പിച്ച ഹാപ്ലോടൈപ്പിന് സമാനമാണെന്ന് കണ്ടെത്തി. ഹാപ്ലോഗ് ഗ്രൂപ്പ് I1 ൻ്റെ 968 111 മാർക്കർ ഹാപ്ലോടൈപ്പുകളുടെ ലിസ്റ്റ് അനുസരിച്ച് 57-60, 110, 131, 132, 153-157, 660, 744, 745 എന്നീ നമ്പറുകളുള്ള ഹാപ്ലോടൈപ്പുകളാണ് ഇവ. അടുത്തതായി, ഒരു പ്രൊഫഷണൽ കമ്പ്യൂട്ടർ പ്രോഗ്രാം (PHYLIP) ഉപയോഗിച്ച് എല്ലാ 968 111-മാർക്കർ ഹാപ്ലോടൈപ്പുകളിൽ നിന്നും ഒരു മരം നിർമ്മിച്ചു, കൂടാതെ ഏറ്റവും ബന്ധപ്പെട്ട (ആദ്യത്തെ 12 മാർക്കറുകൾ അനുസരിച്ച്) ഹാപ്ലോടൈപ്പുകൾ തിരിച്ചറിഞ്ഞു. തത്ഫലമായുണ്ടാകുന്ന വൃക്ഷം ചിത്രീകരണ ആവശ്യങ്ങൾക്കായി ചുവടെ കാണിച്ചിരിക്കുന്നു. ഇത് നല്ല സമമിതി കാണിക്കുന്നു, ഇത് മുഴുവൻ വൃക്ഷവും ഒരു പൊതു പൂർവ്വികനിൽ നിന്നാണ് വരുന്നതെന്ന് സൂചിപ്പിക്കുന്നു. എപ്പോൾ ജീവിച്ചിരുന്നു എന്നതും കാണിക്കും. വിവരങ്ങൾക്ക്, മുഴുവൻ വൃക്ഷത്തിൻ്റെയും അടിസ്ഥാന (പൂർവിക) ഹാപ്ലോടൈപ്പ് ഇപ്രകാരമാണ് 13 22 14 10 13 14 11 14 11 12 11 28 – 15 8 9 8 11 23 16 20 28 12 14 15 16 – 10 10 19 21 14 14 16 20 35 37 12 10 – 11 8 15 15 8 11 10 8 9 9 12 23 25 15 10 12 12 16 8 13 25 20 13 13 11 12 11 11 12 11 – 32 12 8 17 12 24 27 19 11 12 12 13 11 9 11 11 10 12 12 31 11 13 21 16 11 10 24 15 19 11 24 17 13 15 25 12 22 18 12 14 18 9 12 11 ആദ്യത്തെ 12 മാർക്കറുകളിൽ അവതരിപ്പിച്ച ഹാപ്ലോടൈപ്പ് അതിൽ നിന്ന് ഒരു മ്യൂട്ടേഷൻ കൊണ്ട് വ്യത്യാസപ്പെട്ടിരിക്കുന്നു (തുടർച്ചയായ അഞ്ചാമത്തെ മ്യൂട്ടേഷൻ, ഹൈലൈറ്റ് ചെയ്തിരിക്കുന്നു). മൊത്തത്തിൽ, എല്ലാ 968 ഹാപ്ലോടൈപ്പുകളും ആദ്യത്തെ 67 മാർക്കർ പാനലിൽ നിന്ന് 14,891 മ്യൂട്ടേഷനുകളും എല്ലാ 111 മാർക്കറുകളിലും 24,990 മ്യൂട്ടേഷനുകളും കാണിക്കുന്നു. സാധാരണ പൂർവ്വികൻ ജീവിച്ചിരുന്നത് 14891/968/0.12 = 128 → 147 പരമ്പരാഗത തലമുറകൾ (25 വർഷം വീതം), അല്ലെങ്കിൽ 24990/968/0.198 = 130 → 149 പരമ്പരാഗത തലമുറകൾ (0.12, 0.7 19 എന്നിവയ്ക്ക് 1 മ്യൂട്ടേഷൻ നിരക്ക്-1 മ്യൂട്ടേഷൻ നിരക്ക് 0.19 ആണ്. മാർക്കർ ഹാപ്ലോടൈപ്പുകൾ, അമ്പ് - ആവർത്തിച്ചുള്ള മ്യൂട്ടേഷനുകൾക്കുള്ള പട്ടിക തിരുത്തൽ), അല്ലെങ്കിൽ യഥാക്രമം 3675 ± 370, 3725 ± 375 വർഷങ്ങൾക്ക് മുമ്പ് പൊതു പൂർവ്വികൻ. 67-ഉം 111-ഉം മാർക്കർ ഹാപ്ലോടൈപ്പുകൾ ഏതാണ്ട് സമാനമായ ഫലങ്ങൾ നൽകുന്നു, 3700 വർഷത്തെ പശ്ചാത്തലത്തിൽ 50 വർഷത്തെ വ്യത്യാസം മാത്രം.
ഒരേ 12 മാർക്കറുകൾ/അലീലുകൾ അടങ്ങിയ ഈ ഏഴ് ഹാപ്ലോടൈപ്പുകൾ മരത്തിൻ്റെ മൂന്ന് വ്യത്യസ്ത ശാഖകളിലാണ് സ്ഥിതി ചെയ്യുന്നത്. തീർച്ചയായും, അവതരിപ്പിച്ച ഹാപ്ലോടൈപ്പിന് 25, 37 അല്ലെങ്കിൽ അതിലധികമോ മാർക്കറുകൾ ഉണ്ടെങ്കിൽ, തീർച്ചയായും ഒരു ശാഖ മാത്രമേ ഉയർന്നുവരുമായിരുന്നു, എന്നാൽ അത് എന്താണ്. തത്വത്തിൽ, ഇത് ഞങ്ങളെ വളരെയധികം തടസ്സപ്പെടുത്തില്ല. ഈ ശാഖകൾ ഇപ്രകാരമാണ്:
13 22 14 10 13
14 11 14 11 12 11 28 — 1600± 240 വർഷം മുമ്പ് (അവതരിപ്പിച്ച ഹാപ്ലോടൈപ്പിൽ നിന്നുള്ള മൂന്ന് മ്യൂട്ടേഷനുകൾ ഹൈലൈറ്റ് ചെയ്തിട്ടുണ്ട്). അവതരിപ്പിച്ച ഹാപ്ലോടൈപ്പ്, തത്വത്തിൽ, ഈ ശാഖകളിലൊന്നിൽ ആകാം; അതിൻ്റെ 12 മാർക്കറുകൾ മാത്രമേ നാല് ശാഖകളിൽ ഒന്നിനെയും ഒഴിവാക്കാൻ ഞങ്ങളെ അനുവദിക്കുന്നില്ല. ഈ നാല് ശാഖകളുടെ പൊതു പൂർവ്വികർ പൂർവ്വിക ഹാപ്ലോടൈപ്പുകളുടെ വലതുവശത്ത് സൂചിപ്പിച്ചതുപോലെ ജീവിച്ചിരുന്നു. മേൽപ്പറഞ്ഞ സംഖ്യകളുമായി ബന്ധപ്പെട്ട ഹാപ്ലോടൈപ്പുകൾ ഏതൊക്കെ ഉപക്ലേഡുകളുടേതാണെന്നും അതിൻ്റെ ആദ്യ 12 മാർക്കറുകൾ പൂർണ്ണമായും ആവർത്തിക്കുന്നതായും നോക്കാം. ഈ മെറ്റീരിയലിൻ്റെ അവസാനം, ഈ നാല് വ്യക്തികൾക്കുള്ള ജീനോമിക് ഡാറ്റ നൽകിയിരിക്കുന്നു, അതിൽ M253 ൻ്റെ താഴെയുള്ള SNP-കൾ രേഖപ്പെടുത്തിയിട്ടുണ്ട്. മിക്കവാറും, ID00020 ന് സമാന സ്നിപ്പുകൾ ഉണ്ടായിരിക്കും. പൊതുവേ, ഈ ഹാപ്ലോടൈപ്പുകളിൽ ഭൂരിഭാഗത്തിനും SNP-കൾ ഉണ്ട്: I1-M253 > DF29 > CTS6364 > M227, മറ്റൊരു ശാഖയിൽ I1-M253 > DF29 > Z58 > Z138. ID00020 സ്നിപ്പുകൾ ആദ്യത്തെ അല്ലെങ്കിൽ രണ്ടാമത്തെ ശൃംഖലയിൽ ഉൾപ്പെട്ടിരിക്കണം. അവയെല്ലാം മോസ്കോ ലബോറട്ടറി ഓഫ് ഡിഎൻഎ വംശാവലിയിൽ നിർണ്ണയിക്കപ്പെടും. അവതരിപ്പിച്ച ഹാപ്ലോടൈപ്പിലേക്കുള്ള ചരിത്ര പാത ഏകദേശം ഇപ്രകാരമാണ്. ഏകദേശം 43 ആയിരം വർഷങ്ങൾക്ക് മുമ്പാണ് ഹാപ്ലോഗ് ഗ്രൂപ്പ് I രൂപീകരിച്ചത് (സംയോജിത ഹാപ്ലോഗ് ഗ്രൂപ്പ് IJ ൽ നിന്ന്), ഏകദേശം 28 ആയിരം വർഷങ്ങൾക്ക് മുമ്പ് ഇത് ഹാപ്ലോഗ് ഗ്രൂപ്പുകളായി I1, I2 എന്നിവയിലേക്ക് വ്യതിചലിച്ചു (“വ്യതിചലിച്ചത്” എന്നത് ഇവിടെ ഒരു ആപേക്ഷിക പദമാണെങ്കിലും അവ വ്യതിചലിച്ചില്ല, ഇവ സ്വതന്ത്ര സംഭവങ്ങളായിരുന്നു) , ആദ്യത്തേതിൽ നിന്ന് (രണ്ടാമത്തേത് പോലെ) കാലക്രമേണ നിരവധി ഉപവിഭാഗങ്ങൾ രൂപപ്പെട്ടു. സ്വീഡനിലും മധ്യ യൂറോപ്പിലും 7,000 വർഷം പഴക്കമുള്ള ഫോസിലൈസ് ചെയ്ത അസ്ഥികൂട അവശിഷ്ടങ്ങളിൽ നിന്നാണ് ഹാപ്ലോഗ് ഗ്രൂപ്പ് I കണ്ടെത്തിയത്. 4600-4000 വർഷങ്ങൾക്ക് മുമ്പ്, എർബിൻസ് യൂറോപ്പിൽ കുടിയേറുന്ന സമയത്ത്, മധ്യ യൂറോപ്പിൽ നിന്നുള്ള ഹാപ്ലോഗ് ഗ്രൂപ്പ് R1b, ഹാപ്ലോഗ് ഗ്രൂപ്പ് I1 എന്നിവയുടെ വാഹകർ ഏതാണ്ട് പൂർണ്ണമായും അപ്രത്യക്ഷമായി, പക്ഷേ ഒരാൾ അതിജീവിച്ചു, ഇതിൻ്റെ ഫലമായി ഹാപ്ലോഗ് ഗ്രൂപ്പ് I1 ൻ്റെ ആധുനിക വാഹകർക്ക് താരതമ്യേന ഏകദേശം 3,700 വർഷങ്ങൾക്ക് മുമ്പ്, അതായത് ബിസി 2-ആം സഹസ്രാബ്ദത്തിൻ്റെ ആദ്യ പകുതിയിൽ, അതിജീവിച്ച പിൻഗാമികൾക്ക് ഇത് കാരണമായ സമീപകാല പൊതു പൂർവ്വികർ. ഹാപ്ലോഗ് ഗ്രൂപ്പ് I1 ൽ ഒരു പൊതു പൂർവ്വികൻ മാത്രമുള്ളതിനാൽ, ഹാപ്ലോഗ് ഗ്രൂപ്പിൻ്റെ ഹാപ്ലോടൈപ്പ് വൃക്ഷം വളരെ സമമിതിയാണ്. ഇത് വളരെ അപൂർവ്വമായി സംഭവിക്കുന്നു. ID00020 ഈ പൊതു പൂർവ്വികൻ്റെ പിൻഗാമിയാണ്. അപേക്ഷ. ID00020 ബ്രാഞ്ചിൽ നിന്നുള്ള ഹാപ്ലോടൈപ്പുകളുടെ വാഹകരിൽ കാണപ്പെടുന്ന SNP-കളുടെ ഘടനയെക്കുറിച്ചുള്ള വിശദമായ ജീനോമിക് ഡാറ്റ ഞങ്ങൾ ഇവിടെ നൽകുന്നു. ടെർമിനൽ സ്നിപ്പുകൾ ഒഴികെ ഇതിന് ഏതാണ്ട് സമാനമായിരിക്കണം. എന്നാൽ മുകളിൽ പറഞ്ഞ പല കേസുകളിലെയും പോലെ അവ സമാനമാണ്. M223-ലും താഴെയുമുള്ള SNP-കൾ ഹൈലൈറ്റ് ചെയ്തിരിക്കുന്നു: കിറ്റ് നമ്പർ. 38184 (മത്തായി ഹാമിൽട്ടൺ ബുച്ചർ, അമേരിക്കൻ ഇന്ത്യക്കാരൻ, എന്നാൽ മുകളിലെ ശാഖയിൽ ഒരു യൂറോപ്യൻ, ഹാപ്ലോടൈപ്പ് 110-ൽ നിന്ന് Y-DNA ലഭിച്ചു): DF29+, M227+, CTS10058+, CTS10140+, CTS10338+, CTS10834+, CTS11036+, CTS11042+, CTS11441+, CTS11526+, CTS11552+, CTS11552+, CTS11775+, CTS11775 CTS2193+, CTS2375+, CTS2514+, CTS2524+, CTS2644+, CTS2738+, CTS3517+, CTS3654+, CTS3843+, CTS4088+, CTS4130+ , CTS4295+, CTS4437+, CTS4848+, CTS4982+, CTS5167+, CTS5408+, CTS5513+, CTS565+, CTS5650+, CTS5705+, CTS+571+, CTS1783+, CTS5783+, C89 93+, CTS6140+, CTS6221+, CTS623+, CTS6265+, CTS6395+, CTS641+, CTS6629+, CTS674+ , CTS6932+ , CTS7267+, CTS7329+, CTS7831+, CTS7949+, CTS8333+, CTS8345+, CTS8394+, CTS8420+, CTS8716+, CTS88+, CTS2+CTS929+, CTS2+CTS929, 09+, F3692 +, F719+, L118+, L121+, L123+, L124+, L125+, L132+, L15+, L16+ , L187+, L350+, L403+, L41+, L468+, L470+, L498+, L509+, L574+, L575+, L578+, L740+, L748+, L75+, L750, L750, L750, L751 759+, L772 +, L80+, L81+ , M168+, M170+ , M213+, M227+, M235+, M253+, M258+, M294+, M299+, M307+, M89+, M94+, P123+, P124+, P125+, P126+, P127+, P129+, P130+, P133+, P134+, P13+P,13+ P,130, P136+ , P141+, P143+, P145+, P146+ , P148+, P149+, P151+, P157+, P158+, P159+, P160+, P161+, P163+, P166+, P187+, P212+, P30+, P316+, PAGES00020+0, PAGES00020+0, PAGES000200, PAGES2050 F2608+, PF2611+, PF2615+, PF2747+, PF2748+, PF2749+, PF2770+ , PF3562+, PF3574+, PF3639+, PF3640+, PF3641+, PF3660+, PF3666+, PF3672+, PF3675+, PF3677+, PF3780+, PF3+804, PF3800, PF3780+, PF3800, 3814+, PF3815+, PF3819+, PF3822+, PF3836+, PF3837+, PF6464+, PF6469+, PF6470+ , PF6477+ , PF6479+, PF6520+, V241+, V250+, YSC0000207+, YSC0000227+, YSC0000256+, YSC0000272+, YSC0000272+, YSC0000281+0, YSC0000200+1, YSC0000281+0, YSC000281+0, YSC000281+0, YSC000281+0 M72-, Z58-, Z63-, Z17694-, S2348-, S4442-, M3453- കിറ്റ് നമ്പർ. 38737 (ഗ്രേറ്റ്ഹൗസ്, ജർമ്മനി, ഹാപ്ലോടൈപ്പ് 744): CTS10058+, CTS10140+, CTS10338+, CTS10362+, CTS109+, CTS11036+, CTS11042+, CTS11318+, CTS11318+, CTS11318+, CTS11318+, CTS11351, CTS1,55 15 75+, CTS11726+, CTS11775+, CTS11783+, CTS11950+, CTS125+, CTS12632+, CTS1393+, CTS1996+, CTS2193+, CTS2375+, CTS2514+, CTS2524+, CTS2536+, CTS2644+, CTS2738+, CTS3331+, CTS3431+, CTS+3517+, CTS3+36+ 68+, CTS39 96+, CTS4088+, CTS4130+, CTS4295+, CTS4364+, CTS4368+, CTS4443+, CTS4740+, CTS4848+, CTS4982+, CTS5167+, CTS5318+, CTS5408+, CTS5457+, CTS5513+, CTS5532+, CTS565+, CTS5650+, CTS5705+, CTS571+, CTS171+, CTS18+ +, CTS6135+, CTS6140 +, CTS6221+, CTS6265+, CTS6383+, CTS6395+, CTS641+, CTS6629+, CTS674+ , CTS6800+, CTS6907+, CTS6932+, CTS7267+, CTS7329+, CTS7502+, CTS7831+, CTS7922+, CTS7933+, CTS7949+, CTS8243+, CTS2+ CTS34, CTS2+CTS38, CTS2, CTS38 8716+, CTS88+, CTS8876 +, CTS8980+, CTS9240+, CTS9264+, CTS9288+, CTS9828+, F1046+, F1209+, F1302+, F1320+, F1329+, F1450+, F1460+, F1704+, F1714+, F1753+, F1767+, F2048+, F2075+, F2142+, F2142+, F2132+, F21332+, F213256 +, F2408+, F2587+, F2688 +, F2710+, F2794+, F2837+, F2985+ , F2993+, F3033+, F3111+, F3136+, F3335+, F3368+, F3402+, F3556+, F3692+, F4188+, F719+, F922+, L118+, L12+, L12+1, L12+1, L12+1, L12+1, L124+, , L16+, L187+, L350+, L403+ , L468+, L470+ , L498+, L509+, L574+, L575+, L578+, L740+, L748+, L75+, L750+, L751+, L755+, L756+, L758+, L759+, L772+, L759+, L772+, L8+, M1, L8+, M72 M253+ Z138+, Z139+, M170+, M258+, M307+, P19+, P30+, P38+, P109-, P259-, P37-, M161-, M72-, M26-, M21-, M223-, M227- കിറ്റ് നമ്പർ. 152077 (ജെൻസ് പീറ്റർ നീൽസൺ, ഡെൻമാർക്ക്, ഹാപ്ലോടൈപ്പ് 745): CTS10058+, CTS10140+, CTS10338+, CTS10362+, CTS109+, CTS11036+, CTS11042+, CTS14+CTS14, CTS1356 +, C TS11575+, CTS11726+, CTS11775+, CTS11783+, CTS11950+, CTS125+, CTS12632+, CTS1393+ , CTS1996+, CTS2193+, CTS2375+, CTS2514+, CTS2524+, CTS2536+, CTS2644+, CTS2738+, CTS3331+, CTS3431+, CTS3517+, CTS3+ CTS36, CTS3+CTS36 38 68+, CTS3996+, CTS4088+, CTS4130+, CTS4295+, CTS4364+, CTS4368+, CTS4443+, CTS4740+ , CTS4848+, CTS4982+, CTS5167+, CTS5318+, CTS5408+, CTS5457+, CTS5513+, CTS5532+, CTS565+, CTS5650+, CTS+5705+, CTS+571+89, CTS571+89, 93+, CTS6135 +, CTS6140+, CTS6221+, CTS6265+, CTS6383+, CTS6395+, CTS641+, CTS6629+, CTS674+, CTS6800+, CTS6907+, CTS6932+, CTS7267+, CTS7329+, CTS7502+, CTS7831+, CTS7922+, CTS7933+, CTS7949+, CTS4+83+, CTS3 C8243+, C89 0+, CTS8716+, CTS88 +, CTS8876+, CTS8980+, CTS9240+, CTS9264+, CTS9288+, CTS9828+, F1046+ , F1209+, F1302+, F1320+, F1329+, F1450+, F1460+, F1704+, F1714+, F1753+, F1767+, F2048+, F2075+, F2142+, F2236+, F22362+, F2245+, F22465 02+, F2408+, F2587 +, F2688+, F2710+, F2794+, F2837+, F2985+, F2993+, F3033+, F3111+, F3136+, F3335+, F3368+, F3402+, F3556+, F3692+, F4188+, F719+, F922+, L12+, L12+, L12+, L12, L15 L157+, L16+, L187+, L350 +, L403+, L468+ , L470+, L498+, L509+, L574+, L575+, L578+, L740+, L748+, L75+, L750+, L751+, L755+, L756+, L758+, L759+, L759+, M759+, L7072+1, L7072,11 +, M253+, M294+, M307+, M42+, M450+, M89+, M94+, P123+, P124+, P126+, P127+, P130+, P135+, P136+, P138+, P14+, P141+, P141+, P141+, P145, P145, P145 P159+, P160+, P166+, P187+ , P30+, P305+, PAGES00026+, PAGES00081+, PAGES00123+, PF1016+, PF1029+, PF1031+, PF1040+, PF1046+, PF1061+, PF1061+, PF1091+, PF1091, PF1091, PF1091 1269+, PF1276+, PF192+, PF210+, PF212+, PF223+, PF234+, PF258+, PF2591+ , PF2593+ , PF2599+, PF2608+, PF2611+, PF2615+, PF2624+, PF263+, PF2643+, PF272+, PF2745+, PF2747+, PF2748+, PF2748+, PF2748, PF2748, 2+, PF316+ , PF325+, PF342+, PF3515+, PF3517+, PF3518+, PF3534+, PF3560+, PF3561+ , PF3562+ , PF3574+, PF3578+, PF3586+, PF3588+, PF3590+, PF3594+, PF3596+, PF3600+, PF3604+, PF3605+, PF3605+, PF3605+, PF6+P3611, PF3611, PF3611 F3625+, PF 3639+, PF3640+, PF3641+, PF3642+, PF3654+, PF3660+, PF3666+, PF3672+ , PF3675+, PF3677+ , PF3686+, PF3694+, PF3780+, PF3800+, PF3804+, PF3806+, PF3807+, PF3809+, PF3811+, PF3811+, PF3811+, PF392,81, PF3814 F3836+, PF3837+, PF 500+, PF667+, PF719+, PF725+, PF779+, PF796+, PF803+ , PF815+, PF821+, PF840+ , PF844+, PF892+, PF937+, PF951+, PF954+, PF970+, V168+, V186+, V189+, V205+, V2+C, V220+, V220+, V220+5 56+, YSC0000256+, YSC00002 57+, YSC0000259+, YSC0000260+, YSC0000264+ , YSC0000265+, YSC0000267+, YSC0000272+, YSC0000280+ , YSC0000281+, YSC0000298+, YSC0000299+, YSC0000300+, YSC000300+, YSC00000 Z138+, Z139+, M170+, M258+, M307+, P19+, P30+, P38+, P109-, P259-, P37-, M161-, M72-, M26-, M21-, M223-, M227- കിറ്റ് N10060 (റോബർട്ട് വേഡ്, ഇംഗ്ലണ്ട്, ഹാപ്ലോടൈപ്പ് 57): M170+, M253+, M258+, M307+, P19+, P30+, P38+, P109-, P259-, P37-, M161-, M72-, M26-, M21-, M223-, M227- ഹാപ്ലോഗ് ഗ്രൂപ്പ് I2a രണ്ട് പേർക്ക് ഈ ഹാപ്ലോഗ് ഗ്രൂപ്പ് ഉണ്ടായിരുന്നു, ഒന്ന് സബ്ക്ലേഡ് I2a1-L147.2, രണ്ടാമത്തേത് - I2a2-M223, യഥാക്രമം 67-ഉം 111-ഉം മാർക്കർ ഹാപ്ലോടൈപ്പുകൾ. KLIN ID00004 ഒരു 67-മാർക്കർ ഹാപ്ലോടൈപ്പ് അവതരിപ്പിച്ചിരിക്കുന്നു, ഹാപ്ലോഗ് ഗ്രൂപ്പ് I2a റിപ്പോർട്ട് ചെയ്യുന്നു. 13 24 17 11 14 15 11 13 12 13 11 30 – 17 8 10 11 11 25 15 20 0 12 14 15 15 – 10 10 20 21 15 12 19 17 35 35 11 10 – 11 8 15 15 7 12 10 8 11 9 12 22 22 16 10 12 12 12 7 10 30 21 13 14 10 13 11 11 12 9 ഈ ഹാപ്ലോഗ് ഗ്രൂപ്പ് കിഴക്കൻ യൂറോപ്പിലുടനീളം, അതായത് ഗ്രീസ്, സെർബിയ, ബോസ്നിയ-ഹെർസഗോവിന, മാസിഡോണിയ, ചെക്ക് റിപ്പബ്ലിക്, സ്ലൊവാക്യ, പോളണ്ട്, ബൾഗേറിയ, ബെലാറസ്, റഷ്യ, ഉക്രെയ്ൻ, ഹംഗറി, അതുപോലെ ജർമ്മനി, റൊമാനിയ, എന്നിവിടങ്ങളിൽ വിതരണം ചെയ്യപ്പെടുന്നു. ഇറ്റലി, കൂടാതെ പരീക്ഷിച്ചവരിൽ ബഹുഭൂരിപക്ഷത്തിനും L147.2 എന്നും അറിയപ്പെടുന്ന I2a-CTS10228 SNP ഉണ്ടെന്ന് കണ്ടെത്തി (ചുവടെയുള്ള ISOGG ഡയഗ്രം കാണുക; YFull കമ്പനി അനുസരിച്ച് SNP രൂപീകരണത്തിൻ്റെ ഡേറ്റിംഗ് വലതുവശത്താണ്). ഇതൊരു സാധാരണ കിഴക്കൻ യൂറോപ്യൻ സ്നിപ്പാണ്. ഹാപ്ലോടൈപ്പിൻ്റെ രൂപം അനുസരിച്ച്, ഉപഭോക്താവിനും അത് ഉണ്ടെന്ന് വ്യക്തമാണ്. ചോദ്യം ഇതാണ്: ഹാപ്ലോടൈപ്പിൻ്റെ തരത്തെ അടിസ്ഥാനമാക്കി നമുക്ക് ചില താഴ്ന്ന നിലയിലുള്ള എസ്എൻപികൾ പ്രവചിക്കാൻ കഴിയുമോ?
ഡാറ്റാബേസിൽ (FTDNA I2a പ്രൊജക്റ്റ്) 67-മാർക്കർ ഫോർമാറ്റിൽ 245 ഹാപ്ലോടൈപ്പുകൾ അടങ്ങിയിരിക്കുന്നു. ഈ ഹാപ്ലോടൈപ്പുകളിൽ നിന്നെല്ലാം നിർമ്മിച്ച മരത്തിൽ, ഹാപ്ലോടൈപ്പ് ID00004 വലതുവശത്താണ് (എക്സ് ഉപയോഗിച്ച് അടയാളപ്പെടുത്തിയത്), ഇത് ബന്ധപ്പെട്ട വലിയ ശാഖയിൽ പെടുന്നു, അത് ഏറ്റവുമധികം ഉൾക്കൊള്ളുന്നു. വലത്, ഒതുക്കമുള്ള ശാഖ വൃക്ഷം, ഒതുക്കമുള്ള ശാഖകൾ അയഞ്ഞ ശാഖകളുമായി താരതമ്യപ്പെടുത്തുമ്പോൾ താരതമ്യേന ഇളം ശാഖ എന്നാണ് അർത്ഥമാക്കുന്നത്. മറ്റൊരു തരത്തിൽ പറഞ്ഞാൽ, ഒതുക്കമുള്ള ശാഖ അതിൻ്റെ താരതമ്യേന സമീപകാല ഉത്ഭവത്തെ സൂചിപ്പിക്കുന്നു. ഈ ശാഖയ്ക്ക് ഏകദേശം 2300 വർഷം പഴക്കമുണ്ട്, അതായത്, ഇത് രൂപപ്പെട്ടുവെന്ന് കണക്കുകൂട്ടലുകൾ കാണിക്കുന്നു. അവസാന യുഗത്തിൻ്റെ അവസാനത്തിൽ, ഈ ശാഖയുടെ മിക്കവാറും എല്ലാ ഹാപ്ലോടൈപ്പുകളും CTS10228/L147.2 സ്നിപ്പുമായി ബന്ധപ്പെട്ടിരിക്കുന്നു, പക്ഷേ ഇതിന് അവരോഹണ സ്നിപ്പുകളും ഉണ്ട്, ഉദാഹരണത്തിന് L147.2 >YP204/S17250 >Z16971 മരത്തിൽ 67 മാർക്കർ ഹാപ്ലോടൈപ്പുകളിൽ വേർതിരിക്കാത്തവ. കൂടുതൽ കൃത്യമായി പറഞ്ഞാൽ, ഒരുപക്ഷേ അവർക്ക് ബ്രാഞ്ചിൻ്റെ വിവിധ ഭാഗങ്ങളിലേക്ക് വ്യതിചലിച്ചേക്കാം, എന്നാൽ പ്രോജക്റ്റിലെ വളരെ കുറച്ച് ഹാപ്ലോടൈപ്പ് കാരിയറുകളാണ് അത്തരം ആഴത്തിലുള്ള SNP-കളെ തിരിച്ചറിഞ്ഞത്. ഡിഎൻഎ വംശാവലിയുടെ മോസ്കോ ലബോറട്ടറി അവ നിർണ്ണയിക്കും.
111 മാർക്കർ മരത്തിൽ പ്രത്യേക സ്നിപ്പുകൾക്ക് കാരണമായേക്കാവുന്ന ഉച്ചരിച്ച ശാഖകളൊന്നുമില്ലെന്ന് ഞങ്ങൾ കാണുന്നു. 67-മാർക്കർ ഫോർമാറ്റിലുള്ള ശാഖയുടെ അടിസ്ഥാന ഹാപ്ലോടൈപ്പ്: 13 24 16 11 14 15 11 13 13 13 11 31 – 17 8 10 11 11 25 15 20 32 12 14 15 15 – 10 10 21 21 15 12 18 18 34 35 11 10 – 11 8 15 15 7 12 10 8 11 9 12 22 22 16 10 12 12 12 7 10 30 21 13 14 10 13 11 11 12 9 (അവതരിപ്പിച്ച ഹാപ്ലോടൈപ്പിൽ നിന്നുള്ള 8 മ്യൂട്ടേഷനുകൾ തിരിച്ചറിഞ്ഞു). ഇതിനർത്ഥം 8/0.12 = 67 → 72 പരമ്പരാഗത തലമുറകൾ, അതായത് ഏകദേശം 1800 വർഷം കൊണ്ട് പൂർവ്വികനിൽ നിന്ന് ഹാപ്ലോടൈപ്പ് നീക്കം ചെയ്യപ്പെടുന്നു എന്നാണ്. രണ്ട് ഹാപ്ലോടൈപ്പുകൾക്ക് കൃത്യമായ കണക്കുകൂട്ടലുകൾ ഇല്ലാത്തതിനാൽ ഇത് തീർച്ചയായും ഏകദേശം ആണ് - അവതരിപ്പിച്ച ഹാപ്ലോടൈപ്പിലെ ഒരു ക്രമരഹിതമായ മ്യൂട്ടേഷൻ കണക്കാക്കിയ ദൂരത്തെ 200 വർഷത്തിലേറെയായി മാറ്റും. എന്തായാലും, ഉപഭോക്താവിൻ്റെ പൂർവ്വികർ ID00004 ഡിഎൻഎ ലൈൻ ഈ ശാഖയുടെ പൊതു പൂർവ്വികനിൽ നിന്ന് നേരിട്ട് കണ്ടെത്തിയതായി വ്യക്തമാണ്. ഏകദേശം 2300 വർഷം പഴക്കമുള്ള ഹാപ്ലോഗ് ഗ്രൂപ്പ്/സബ്ക്ലേഡ് I2a-L147.2 ൻ്റെ ചരിത്രം റീഫോർമാറ്റിൽ ആവർത്തിച്ച് ചർച്ച ചെയ്യപ്പെട്ടിട്ടുണ്ട്. ഹാപ്ലോഗ് ഗ്രൂപ്പ് I2a മധ്യ യൂറോപ്പിലെ ശ്മശാനങ്ങളിൽ 7000-5000 വർഷങ്ങൾക്ക് മുമ്പുള്ള തീയതികളിൽ കണ്ടെത്തി, പിന്നീട് ഏകദേശം 4500 വർഷങ്ങൾക്ക് മുമ്പ് അത് മധ്യ യൂറോപ്പിൽ നിന്ന് അപ്രത്യക്ഷമാവുകയും രണ്ട് ശാഖകളായി വിഭജിക്കുകയും ചെയ്തു. ഒന്ന് (കൂടുതൽ കൃത്യമായി പറഞ്ഞാൽ, മുകളിലുള്ള ആദ്യ ഡയഗ്രം കാണുക) ബ്രിട്ടീഷ് ദ്വീപുകളിലേക്ക് നീങ്ങി (അല്ലെങ്കിൽ പലായനം ചെയ്തു), ഈ ശാഖകളുടെ പൊതു പൂർവ്വികർ ഏകദേശം 4500-5000 വർഷങ്ങൾക്ക് മുമ്പാണ്. ഉപക്ലേഡ് I2a-L147.2 ൻ്റെ മറ്റൊരു ശാഖ യൂറോപ്പിലെ ജനസംഖ്യാ തടസ്സത്തിലൂടെ കടന്നുപോയി, ഏകദേശം 2300 വർഷങ്ങൾക്ക് മുമ്പ് ഡാന്യൂബ്-കാർപാത്തിയൻസിൽ പുനരുജ്ജീവിപ്പിക്കുകയും എഡി ഒന്നാം സഹസ്രാബ്ദത്തിൻ്റെ ആദ്യ പകുതിയിൽ കിഴക്കൻ യൂറോപ്പിലുടനീളം വ്യാപിക്കുകയും ചെയ്തു. ഒരുപക്ഷേ ചരിത്രകാരന്മാരും പുരാവസ്തു ഗവേഷകരും ഇത് "സ്ലാവുകളുടെ ഉത്ഭവം" ആയി തെറ്റിദ്ധരിച്ചിരിക്കാം, പുരാതന വൃത്താന്തങ്ങളിൽ ഇതിനെക്കുറിച്ച് ചർച്ചചെയ്യുന്നു, ഉദാഹരണത്തിന്, പഴയ വർഷങ്ങളുടെ കഥ. PVL ശരിയാണെങ്കിൽ, Haplogroup I2a-L147.2 ൻ്റെ സ്ലാവുകൾ വടക്കൻ അഡ്രിയാറ്റിക് കടലിൻ്റെ വടക്ക് കിഴക്കൻ ആൽപ്സിലെ നോറിക്കിൽ നിന്നാണ് വന്നത്. നോറിക്കിലെ നിവാസികളായ ടൗറിസ്കന്മാർ ബിസി ഒന്നാം സഹസ്രാബ്ദത്തിൻ്റെ രണ്ടാം പകുതി മുതൽ അറിയപ്പെടുന്നു. ID00004 BigY ടെസ്റ്റ് (അല്ലെങ്കിൽ Y ക്രോമസോമിനുള്ള മറ്റൊരു ജീനോമിക് ടെസ്റ്റ്) നടത്തിയിരുന്നെങ്കിൽ, ഫലം ഇനിപ്പറയുന്നവയോട് അടുത്ത് (അല്ലെങ്കിൽ സമാനമായത്) ആയിരിക്കും (I2a-CTS10228/L147.2 സബ്ക്ലേഡിൻ്റെ ഉയർന്ന SNP-കൾ ഇവിടെ ഹൈലൈറ്റ് ചെയ്തിരിക്കുന്നു). ശേഷിക്കുന്ന അടയാളപ്പെടുത്തിയ SNP-കൾ ഇതിലും ഉയർന്ന ഹാപ്ലോടൈപ്പുകളിൽ പെടുന്നു, ഉദാഹരണത്തിന്, M42 - സംയുക്ത ഹാപ്ലോഗ് ഗ്രൂപ്പ് VT; V9, V52, V189 എന്നിവയെല്ലാം സംയോജിത ഹാപ്ലോഗ് ഗ്രൂപ്പ് എസ്.ടി. M89 എന്നത് ഹാപ്ലോഗ് ഗ്രൂപ്പ് എഫ്, എൽ15, എൽ16 എന്നിവ സംയോജിപ്പിച്ച ഹാപ്ലോഗ് ഗ്രൂപ്പ് IJK ആണ്, L41 എന്നത് ഹാപ്ലോഗ് ഗ്രൂപ്പ് I ആണ്. നിങ്ങൾക്ക് കാണാനാകുന്നതുപോലെ, BigY ടെസ്റ്റ് ($599-ന്) ഇവിടെ നൽകിയ ഒരേയൊരു ഉപയോഗപ്രദമായ കാര്യം ഏറ്റവും കുറഞ്ഞ സ്നിപ്പ് I2a-CTS10228-ൻ്റെ തിരിച്ചറിയൽ മാത്രമാണ്. ഇവിടെ കൂടുതൽ സബോർഡിനേറ്റ് സ്നിപ്പുകളൊന്നുമില്ല, ചുവടെയുള്ള പട്ടികയിലെ മുകളിലെ സ്നിപ്പ് Z17855 നെഗറ്റീവ് ആണ് (അവസാനം മുതൽ നാലാമത്തേത്). താഴെ നൽകിയിരിക്കുന്ന സ്നിപ്പുകളുടെ കൂട്ടം ഒരു തുറന്ന ഡാറ്റാബേസിൽ നിന്നാണ് എടുത്തത്; ഇത് പോളിഷ് രാജകുമാരൻ സ്വ്യാറ്റോപോക്ക്-ചെറ്റ്വെർട്ടിൻസ്കിയുടെ പിൻഗാമിയാണ്, ഐതിഹ്യമനുസരിച്ച് - പോളിഷ് റൂറിക്കോവിച്ച്സിൽ നിന്ന്. വഴിയിൽ, റഷ്യൻ സൈന്യത്തിൻ്റെ കേണലായിരുന്ന നെപ്പോളിയൻ യുദ്ധങ്ങളിൽ പങ്കെടുത്ത പ്രിൻസ് ബോറിസ് അൻ്റോനോവിച്ച് ചെറ്റ്വെർട്ടിൻസ്കി (1784-1865) അതേ പോളിഷ് കുടുംബത്തിൽ നിന്നുള്ളയാളാണ്. ഈ സ്നിപ്പുകളിൽ ഭൂരിഭാഗവും, അല്ലെങ്കിലും, ID00004-ൽ കണ്ടെത്തണം. എന്നിരുന്നാലും, ആഴത്തിലുള്ള സബ്ക്ലേഡ് ചെറ്റ്വെർട്ടിൻസ്കിയുടേതിന് സമാനമായി മാറിയാൽ മാത്രം മതി. എന്നാൽ ചെറ്റ്വെർട്ടിൻസ്കിയിൽ L147.2-ന് താഴെയുള്ള ആഴത്തിലുള്ള സ്നിപ്പുകളൊന്നും കണ്ടെത്തിയില്ല എന്നത് രസകരമാണ്; അവസാനത്തെ ടെർമിനൽ CTS10228 ആണ്, അതായത് L147.2. CTS4002+, CTS4039+, CTS4088+, CTS410+, CTS4437+, CTS4848+, CTS4982+, CTS5044+, CTS5375+, CTS5650+, CTS5727+, CTS5884+, C69, CTS5884+, C69 +, CTS6265+, CTS674+, CTS6932+, CTS7175+, CTS7213+, CTS7218+, CTS7329+, CTS7831+, CTS8239+ , CTS8333+, CTS8345+, CTS8420+, CTS8486+, CTS88+, CTS8876+, CTS9240+, CTS9264+, CTS9349+, CTS10058+, CTS10228+, CTS10834+, CTS10936+, CTS11030+, CTS11441+, CTS11768+, CTS1293+, CTS176+, CTS1846+, CTS2193+, CTS2514+, CTS3296+, CTS32931+, CTS3296+, CTS3296+, CTS,256 F3692+, F719+, L132+, L15+, L16+, L178+, L350+, L403+, L41+, L460+, L468+, L470+, L498+, L578+, M42+, M423+, M438+, M89+, M94+, P123+, P124+, P125+, P126+, P127+, P129+, P130+, P133+, P134+, P135+, P136+, P138+, P135+, P136+, P138+, P14+ P,14+ P,14 , P146+, P148+, P149+, P151+ , P157+, P158+, P159+, P160+, P161+, P163+, P166+, P187+, P212+, P316+, P38+, PAGES00026+, PAGES00081+, PF2591+, PF2591+,P2591+,P2591+,P2591+,P2562,F1,F4 7+, PF2748+, PF2749+, PF2770+, PF3562+, PF3574+, PF3626+ , PF3638+ , PF3639+, PF3640+, PF3641+, PF3644+, PF3651+, PF3652+, PF3657+, PF3658+, PF3660+, PF3664+, PF3664+, PF3664+, PF3666+ PF3666, PF3666 F3672+, PF 3675+, PF3677+, PF3780+, PF3800+, PF3801+, PF3802+, PF3804+, PF3809+ , PF3811+, PF3812+ , PF3814+, PF3815+, PF3819+, PF3820+, PF3822+, PF3836+, PF3837+, PF3849+, PF3876+, PF3876+, PF3876+6+ PF3946, PF6965 F6470+, PF6477+, PF 6479+, PF6520+, L621+, L68+, L748+, L751+, L755+, L756+, L758+, L772+, M168+, M170+, M213+, M235+, M258+, M294+, M299+, V241+, V250+, YSC0000207+, YSC0000227+, YSC0000256+, YSC0000272+, M423+, Z421, Z3 -, Z2109-, M3453-, S2348-, S4442- , P61-, L584-, CTS3802-, DF41- നമ്മൾ റൂറിക്കോവിച്ചുകളെക്കുറിച്ചാണ് സംസാരിക്കുന്നത് എന്നതിനാൽ, റഷ്യൻ അസംബ്ലി ഓഫ് നോബിലിറ്റിയിലെ രാജകുമാരന്മാരുടെ ഗ്രൂപ്പിന് ഹാപ്ലോഗ് ഗ്രൂപ്പ് N1c1 ഉണ്ടെന്നും ചെറ്റ്വെർട്ടിൻസ്കിക്ക് (ഉപഭോക്താവിനും) ഹാപ്ലോഗ് ഗ്രൂപ്പ് I2a ഉണ്ടെന്നും നമുക്ക് ഓർക്കാം. റൂറിക്ക്, അത്തരമൊരു സംഗതി നിലവിലുണ്ടെങ്കിൽ, ഒരു ഹാപ്ലോഗ് ഗ്രൂപ്പ് മാത്രമേ ഉണ്ടാകൂ. അതിനാൽ, തങ്ങൾ റൂറിക്കോവിച്ച്മാരാണെന്ന് അവകാശപ്പെടാൻ എല്ലാവർക്കും ഡോക്യുമെൻ്ററി അടിസ്ഥാനമുണ്ടെങ്കിൽ, നിരവധി റൂറിക്കോവിച്ചുകളുടെ ചോദ്യം ഇപ്പോഴും തുറന്നിരിക്കുന്നു. എനിക്കറിയാവുന്നിടത്തോളം, ആർക്കും ഡോക്യുമെൻ്ററി തെളിവുകളൊന്നുമില്ല, പക്ഷേ നാട്ടുരാജ്യങ്ങളുടെ വംശാവലിയെക്കുറിച്ച് വിദഗ്ധരുടെ സുസ്ഥിരമായ അഭിപ്രായങ്ങളുണ്ട്. മാത്രമല്ല, പോളിഷ് വംശശാസ്ത്രജ്ഞർക്ക് ഒരുപക്ഷേ ഒരു അഭിപ്രായമുണ്ട്, റഷ്യൻ വംശശാസ്ത്രജ്ഞർക്ക് വ്യത്യസ്ത അഭിപ്രായമുണ്ട്. അവർ പരസ്പരം കാര്യങ്ങൾ മനസ്സിലാക്കാൻ ശ്രമിക്കുന്നുണ്ടാകാം, പക്ഷേ ഡിഎൻഎ വംശാവലി അവർക്ക് പരിഗണിക്കാൻ പുതിയ ഡാറ്റ നൽകുന്നു. ഒരുപക്ഷേ മോസ്കോ ഡിഎൻഎ വംശാവലി ലബോറട്ടറിക്ക് ID00004-ൻ്റെ താഴത്തെ SNP-കൾ (CTS1028-മായി ബന്ധപ്പെട്ട്) തിരിച്ചറിയാനും അവ ഏതൊക്കെ മേഖലകളിലാണ് പ്രധാനമായും പ്രകടിപ്പിക്കുന്നതെന്ന് മനസ്സിലാക്കാനും കഴിഞ്ഞേക്കും. അപ്പോൾ വ്യക്തിഗത വംശാവലി കൂടുതൽ ആഴത്തിലാക്കാം. KLIN ID00019 111-മാർക്കർ ഹാപ്ലോടൈപ്പ് അവതരിപ്പിച്ചിരിക്കുന്നു, ഹാപ്ലോഗ് ഗ്രൂപ്പ് I2b, സബ്ക്ലേഡ് I2b1, സ്നിപ്പ് M223 എന്നിവ റിപ്പോർട്ട് ചെയ്യപ്പെടുന്നു. 15 23 15 10 15 15 11 14 12 14 12 32 16 8 9 11 11 25 14 20 28 11 14 14 15 11 10 19 21 15 14 18 21 32 40 12 10 11 8 16 16 8 11 10 8 10 9 12 22 22 15 11 12 12 15 9 14 27 21 11 13 12 12 11 12 12 11 32 13 8 15 10 24 27 16 12 11 14 11 12 9 13 11 10 10 12 32 10 12 22 13 11 10 20 15 23 9 24 14 12 14 27 12 21 18 12 15 17 9 12 11 അവതരിപ്പിച്ച ഡാറ്റ 2010 മുതൽ കാലഹരണപ്പെട്ട നാമകരണം ഉപയോഗിക്കുന്നു; അതിനുശേഷം, SNP M223 സബ്ക്ലേഡ് I2a2a ആയി തരംതിരിച്ചിട്ടുണ്ട് (ചുവടെയുള്ള ഡയഗ്രം, ചുരുക്കരൂപത്തിൽ കാണുക). ജെനോമിക് ഡാറ്റ ഉപയോഗിച്ച് YFull കമ്പനി കണക്കാക്കിയ ഏറ്റവും പഴയതും മറ്റ് ചില എസ്എൻപികളുടേയും (ഇവിടെ ഞങ്ങൾക്ക് താൽപ്പര്യമുള്ളവ) ഉത്ഭവത്തിൻ്റെ ഏകദേശ ഡേറ്റിംഗ് (വർഷങ്ങളിൽ, വർഷങ്ങൾക്ക് മുമ്പ്) വലത് കോളം കാണിക്കുന്നു.
മുകളിലെ ലിലാക്ക് നിറം സബ്ക്ലേഡ് I2a-L147.2 സൂചിപ്പിക്കുന്നു, കിഴക്കൻ യൂറോപ്പിലെ ഹാപ്ലോഗ് ഗ്രൂപ്പ് I-ൽ ഏറ്റവും സാധാരണമായത് - ഗ്രീസ് മുതൽ ബാൾട്ടിക് രാജ്യങ്ങൾ വരെ, റഷ്യ, ഉക്രെയ്ൻ, ബെലാറസ്, മുൻ യുഗോസ്ലാവിയയുടെ രാജ്യങ്ങൾ; അവതരിപ്പിച്ച ഹാപ്ലോടൈപ്പായ I2a-M223-ൻ്റെ ഉപവിഭാഗത്തെ ചുവന്ന വര ഹൈലൈറ്റ് ചെയ്യുന്നു. സബ്ക്ലേഡ് തന്നെ പുരാതനമാണ്, ഇതിന് 17,400 വർഷം പഴക്കമുണ്ട്, തീർച്ചയായും ഇതിന് താഴ്ന്ന ഉപക്ലേഡുകൾ ഉണ്ട്, അത് ഈ വ്യാഖ്യാനത്തിൽ തിരിച്ചറിയാൻ ഞങ്ങൾ അടുത്തുവരും. ID00019-ന് ഉണ്ടായിരിക്കാവുന്ന ഉപക്ലേഡുകൾ താഴെ കാണിച്ചിരിക്കുന്നതുപോലെ മഞ്ഞ നിറത്തിൽ ഹൈലൈറ്റ് ചെയ്തിരിക്കുന്നു. FTDNA പ്രോജക്റ്റ് I-ൽ നിന്നുള്ള ഹാപ്ലോഗ് ഗ്രൂപ്പ് I2 ൻ്റെ 111-മാർക്കർ ഹാപ്ലോടൈപ്പുകളുടെ പൊതു വൃക്ഷത്തിൽ അവതരിപ്പിച്ച ഹാപ്ലോടൈപ്പിൻ്റെ സ്ഥാനം നിർണ്ണയിക്കാൻ ശ്രമിക്കാം. ഇത് ചെയ്യുന്നതിന്, ഞങ്ങൾ പ്രോജക്റ്റിൻ്റെ ഹാപ്ലോഗ് ഗ്രൂപ്പ് I2 ൻ്റെ എല്ലാ 63 111-മാർക്കർ ഹാപ്ലോടൈപ്പുകളും എടുക്കും, അവയിൽ അവതരിപ്പിച്ച ഹാപ്ലോടൈപ്പ് ID00019 ചേർക്കുക, കൂടാതെ ഒരു പ്രൊഫഷണൽ കമ്പ്യൂട്ടർ പ്രോഗ്രാം (PHYLIP) ഉപയോഗിച്ച് ഫലമായുണ്ടാകുന്ന 64 ഹാപ്ലോടൈപ്പുകളിൽ നിന്ന് ഒരു മരം നിർമ്മിക്കുകയും ചെയ്യും. ഏതൊക്കെ ഹാപ്ലോടൈപ്പുകൾ, ഏത് ഉപക്ലേഡുകളിൽ നിന്നാണ് ഹാപ്ലോടൈപ്പ് ചുറ്റപ്പെട്ടിരിക്കുന്നതെന്ന് കണ്ടെത്തുക എന്നതാണ് ഈ പരീക്ഷണത്തിൻ്റെ ലക്ഷ്യം. ഹാപ്ലോഗ് ഗ്രൂപ്പ് I2 ൻ്റെ ഈ 111-മാർക്കർ ഹാപ്ലോടൈപ്പുകളുടെ വൃക്ഷം ചുവടെ നൽകിയിരിക്കുന്നു.
അവതരിപ്പിച്ച ഹാപ്ലോടൈപ്പിൻ്റെ സ്ഥാനം, സൂചിക X വഴി വൃക്ഷത്തിൽ സൂചിപ്പിച്ചിരിക്കുന്നു. കാഴ്ചയിൽ സമാനമായ, സാധ്യതയുള്ള ഹാപ്ലോടൈപ്പുകളുടെ, പ്രത്യേകമായി, നന്നായി നിർവചിക്കപ്പെട്ട ശാഖയാണിത്. വിവരങ്ങൾക്ക്, മുഴുവൻ I2 ട്രീയുടെയും അടിസ്ഥാന (പൂർവിക) ഹാപ്ലോടൈപ്പ് ഇപ്രകാരമാണ് 13 24 16 10 13 15 11 13 12 13 11 30 17 8 10 11 11 25 15 20 29 12 14 15 15 10 10 19 20 15 14 17 18 34 36 12 10 11 8 15 16 8 11 10 8 11 9 11 21 22 16 11 12 12 15 8 13 25 21 11 13 11 13 11 12 12 11 30 14 8 15 11 25 27 18 12 11 12 12 12 9 12 11 10 11 12 31 11 12 22 14 11 10 22 15 20 11 23 16 11 15 25 12 22 18 12 14 17 9 12 11 അവതരിപ്പിച്ച ഹാപ്ലോടൈപ്പ് അതിൽ നിന്ന് 70 മ്യൂട്ടേഷനുകളാൽ വ്യത്യാസപ്പെട്ടിരിക്കുന്നു. ഇത് 70/0.198 = 354 → 537 സോപാധിക തലമുറകൾക്ക് തുല്യമാണ്, അല്ലെങ്കിൽ അവയ്ക്കിടയിലുള്ള ഏകദേശം 13425 വർഷങ്ങൾ (0.198 എന്നത് 111-മാർക്കർ ഹാപ്ലോടൈപ്പുകളുടെ മ്യൂട്ടേഷൻ റേറ്റ് സ്ഥിരാങ്കമാണ്, ആവർത്തിച്ചുള്ള മ്യൂട്ടേഷനുകളുടെ തിരുത്തലാണ് അമ്പ്). മറ്റൊരു വിധത്തിൽ പറഞ്ഞാൽ, ഹാപ്ലോഗ് ഗ്രൂപ്പിൻ്റെ പൊതു പൂർവ്വികൻ ID00019-ൽ നിന്ന് വളരെ അകലെയാണ്. എല്ലാ 64 ഹാപ്ലോടൈപ്പുകളിലും അടിസ്ഥാന ഹാപ്ലോടൈപ്പിൽ നിന്ന് 3906 മ്യൂട്ടേഷനുകൾ ഉണ്ട്, ഇത് ഈ ഹാപ്ലോടൈപ്പിൻ്റെ ആധുനിക വാഹകരുടെ പൊതു പൂർവ്വികൻ ജീവിച്ചിരുന്ന സമയവുമായി യോജിക്കുന്നു, ഈ സാഹചര്യത്തിൽ ഇത് 3906/64/0.198 = 308 → 440 സോപാധിക തലമുറകൾ (25 വർഷം വീതം, ഈ കാലയളവിൽ അവ കാലിബ്രേറ്റ് ചെയ്ത മ്യൂട്ടേഷൻ റേറ്റ് സ്ഥിരാങ്കങ്ങളായിരുന്നു), അല്ലെങ്കിൽ 11000± 1100 വർഷങ്ങൾക്ക് മുമ്പ്. സബ്ക്ലേഡ് I2 രൂപീകരിച്ചതിന് ശേഷം അദ്ദേഹം വളരെക്കാലം ജീവിച്ചിരുന്നുവെന്ന് വ്യക്തമാണ് (ഏകദേശം 27,500 വർഷങ്ങൾക്ക് മുമ്പ്, YFull സ്നിപ്പുകളിൽ നിന്ന് നിർണ്ണയിച്ചത്), ഇത് സൂചിപ്പിക്കുന്നത് ഹാപ്ലോഗ് ഗ്രൂപ്പ് I2 ഏകദേശം 11,000 വർഷങ്ങൾക്ക് മുമ്പ്, യൂറോപ്പിലെ ഹിമയുഗവുമായി ബന്ധപ്പെട്ടതാണ്. . ഉപഭോക്താവിൻ്റെയും ഹാപ്ലോഗ് ഗ്രൂപ്പ് I2 ൻ്റെയും പൊതു പൂർവ്വികൻ ഏകദേശം (13425+11000)/2 = 12200 വർഷങ്ങൾക്ക് മുമ്പ്, അതായത് തടസ്സത്തിന് മുമ്പ് ജീവിച്ചിരുന്നു. അവതരിപ്പിച്ച ഹാപ്ലോടൈപ്പ് 39, 44-46, 51, 52 എന്ന നമ്പറുള്ള ഹാപ്ലോടൈപ്പുകളുള്ള അതേ ശാഖയിലാണെന്ന് ഞങ്ങൾ കാണുന്നു. ഈ ശാഖയുടെ അടിസ്ഥാന ഹാപ്ലോടൈപ്പ് 15 23 15 10 15 15 11 13 12 14 12 32 15 8 10 11 11 25 14 20 28 11 14 14 15 11 10 19 21 14 14 17 19 33 39 12 10 11 8 15 16 8 11 10 8 10 9 12 21 22 15 11 12 12 14 9 13 27 20 11 13 12 12 11 12 12 11 31 13 8 15 11 24 27 16 12 11 13 11 13 9 12 11 10 11 12 32 10 12 22 13 11 10 20 15 22 9 23 14 12 14 28 12 21 18 12 15 17 9 12 11 ഇത് അവതരിപ്പിച്ച ഹാപ്ലോടൈപ്പിൽ നിന്ന് 23 മ്യൂട്ടേഷനുകളാൽ (അടയാളപ്പെടുത്തിയത്) വ്യത്യാസപ്പെട്ടിരിക്കുന്നു, അതായത്, 23/0.198 = 116 → 132 പരമ്പരാഗത തലമുറകൾ (25 വർഷം വീതം), അതായത് ഏകദേശം 3300 വർഷം കൊണ്ട് ഈ ശാഖയുടെ പൂർവ്വികനിൽ നിന്ന് ഹാപ്ലോടൈപ്പ് നീക്കംചെയ്യുന്നു. അതേ സമയം, ശാഖയുടെ ഏഴ് ഹാപ്ലോടൈപ്പുകൾക്കും മുകളിൽ കാണിച്ചിരിക്കുന്ന അടിസ്ഥാന ഹാപ്ലോടൈപ്പിൽ നിന്ന് 163 മ്യൂട്ടേഷനുകൾ ഉണ്ട്, അതായത്, ഈ ശാഖയുടെ പൊതു പൂർവ്വികൻ 163/7/0.198 = 118 → 134 പരമ്പരാഗത തലമുറകൾ, അതായത് 3350 ± 425 വർഷം ജീവിച്ചു. മുമ്പ്. ഈ രണ്ട് മൂല്യങ്ങളും ഏതാണ്ട് തികച്ചും യോജിക്കുന്നതായി ഞങ്ങൾ കാണുന്നു (3300, 3350 വർഷം, പ്രത്യേകിച്ച് കണക്കുകൂട്ടൽ പിശകിൻ്റെ പരിധിക്കുള്ളിൽ). ഇതിനർത്ഥം അവതരിപ്പിച്ച ഹാപ്ലോടൈപ്പ് ഏഴ് ഹാപ്ലോടൈപ്പുകളുടെ ഈ ശാഖയുമായി ഏതാണ്ട് തികച്ചും യോജിക്കുന്നു, കൂടാതെ ഹാപ്ലോടൈപ്പിലും അതിൽ നിന്നുള്ള ദൂരത്തിൻ്റെ കാര്യത്തിലും ശാഖയുടെ പൊതു പൂർവ്വികൻ്റെ കണക്കാക്കിയ പ്രായത്തിൻ്റെ കാര്യത്തിലും ഇത് യോജിക്കുന്നു. അങ്ങനെ ഞങ്ങൾ ഹാപ്ലോടൈപ്പ് ID00019 എന്ന ശാഖ കണ്ടെത്തി. 39, 44, 45, 46, 51, 52 എന്നീ നമ്പറുകളുള്ള (ഡിഎൻഎ വംശാവലി അർത്ഥത്തിൽ) ഹാപ്ലോടൈപ്പുകൾ ഏതൊക്കെ ഉപവിഭാഗങ്ങളുടേതാണെന്ന് നോക്കാം. ഹാപ്ലോടൈപ്പ് 39 (ഇംഗ്ലണ്ട്) ഹാപ്ലോടൈപ്പിന് മരത്തിൽ ഏറ്റവും അടുത്താണ്. അദ്ദേഹത്തിന് 3100 വയസ്സുള്ള സ്നിപ്പ് പി 95 ഉണ്ട് (ഈ മെറ്റീരിയലിൻ്റെ തുടക്കത്തിൽ ഡയഗ്രാമിൽ മഞ്ഞ നിറത്തിൽ ഹൈലൈറ്റ് ചെയ്തിട്ടുണ്ട്). ഇത് പ്രായോഗികമായി പരിഗണനയിലുള്ള ബ്രാഞ്ചിൻ്റെ പ്രായവുമായി പൊരുത്തപ്പെടുന്നു, 3350± 425 വർഷം. അടുത്തതായി ഹാപ്ലോടൈപ്പുകൾ 51, 52 (സ്കോട്ട്ലൻഡ്, സ്വീഡൻ) വരുന്നു, അവ ID00019 പോലെ M223-നേക്കാൾ ആഴത്തിലുള്ള SNP-കൾക്കായി ടൈപ്പ് ചെയ്തിട്ടില്ല. എന്നാൽ പ്രോജക്റ്റ് അവയെ Z161, L801, CTS6433 എന്നിവയേക്കാൾ ഉയർന്നതല്ലാത്ത SNP-കളായി നിർവചിക്കുന്നു. ബ്രാഞ്ചിലെ അവസാനത്തെ മൂന്ന് ഹാപ്ലോടൈപ്പുകൾ, 44, 45, 46 (ഇംഗ്ലണ്ട്, പോർച്ചുഗൽ, സ്വിറ്റ്സർലൻഡ്) എന്നിവയ്ക്ക് യഥാക്രമം 2400, 3800, 3800 വർഷങ്ങളുള്ള SNP-കൾ Z79, CTS6433, CTS6433 എന്നിവയുണ്ട്. അതിനാൽ, ID00019 ബ്രാഞ്ച് താരതമ്യേന ചെറുപ്പമാണ്, അതിലെ SNP-കൾ പ്രായവുമായി പൊരുത്തപ്പെടുന്നു, SNP-കളുടെ പ്രായം ഒരു നിശ്ചിത പിശക് ഉപയോഗിച്ചാണ് നിർണ്ണയിക്കുന്നത്. ചുരുക്കത്തിൽ, ID00019 സ്നിപ്പുകൾ വ്യാഖ്യാനത്തിൻ്റെ തുടക്കത്തിൽ സ്നിപ്പ് ഡയഗ്രാമിൽ മഞ്ഞ നിറത്തിൽ സൂചിപ്പിച്ചിരിക്കുന്ന സ്നിപ്പുകളുടെ പരിധിയിലായിരിക്കണം, മിക്കവാറും സ്നിപ്പുകൾ P95, CTS6433, Z79 എന്നിവയിൽ ആയിരിക്കണമെന്ന് നിഗമനം ചെയ്യണം. ഈ എസ്എൻപികളെല്ലാം നിർണ്ണയിക്കാൻ മോസ്കോ ഡിഎൻഎ വംശാവലി ലബോറട്ടറി പദ്ധതിയിടുന്നു. അവതരിപ്പിച്ച ഹാപ്ലോടൈപ്പിലേക്കുള്ള ചരിത്ര പാത ഏകദേശം ഇപ്രകാരമാണ്. ഏകദേശം 43 ആയിരം വർഷങ്ങൾക്ക് മുമ്പാണ് ഹാപ്ലോഗ് ഗ്രൂപ്പ് I രൂപീകരിച്ചത് (ഏകീകൃത ഹാപ്ലോഗ് ഗ്രൂപ്പ് ഐജെയിൽ നിന്ന്), ഏകദേശം 28 ആയിരം വർഷങ്ങൾക്ക് മുമ്പ് ഇത് ഹാപ്ലോഗ് ഗ്രൂപ്പുകളായി ഐ 1, ഐ 2 എന്നിവയിലേക്ക് വ്യതിചലിച്ചു, രണ്ടാമത്തേതിൽ നിന്ന് (അതുപോലെ ആദ്യത്തേതിൽ നിന്നും) നിരവധി താഴ്ന്ന സബ്ക്ലേഡുകൾ രൂപീകരിച്ചു. ഹാപ്ലോഗ് ഗ്രൂപ്പ് I2 ൻ്റെ കൂടുതൽ ചരിത്രം മുൻ വ്യാഖ്യാനങ്ങളിൽ വിവരിച്ചിരിക്കുന്നു; തൽഫലമായി, കിഴക്കൻ യൂറോപ്പിൽ നിന്നുള്ള ഹാപ്ലോഗ് ഗ്രൂപ്പ് I2a യുടെ മിക്ക വാഹകരും L147.2 സബ്ക്ലേഡിൻ്റെ അവരോഹണ ശാഖകളിൽ പെടുന്നു, അവരുടെ പൊതു പൂർവ്വികർ ഏകദേശം 2300 വർഷങ്ങൾക്ക് മുമ്പ് ബിസി ഒന്നാം സഹസ്രാബ്ദത്തിൻ്റെ അവസാനത്തിൽ ജീവിച്ചിരുന്നു. ID00029 മറ്റൊരു സമാന്തര ശാഖയായ M223 ൻ്റെ പ്രതിനിധിയാണ്, എന്നാൽ താരതമ്യേന സമീപകാല ഉപക്ലേഡുകൾ, ഏകദേശം 3300 വർഷങ്ങൾക്ക് മുമ്പ് ബിസി രണ്ടാം സഹസ്രാബ്ദത്തിൻ്റെ രണ്ടാം പകുതിയിൽ ജീവിച്ചിരുന്ന പൊതു പൂർവ്വികൻ. അപേക്ഷ. ID00029 ബ്രാഞ്ചിൽ നിന്നുള്ള ഹാപ്ലോടൈപ്പുകളുടെ വാഹകരിൽ കാണപ്പെടുന്ന SNP-കളുടെ ഘടനയെക്കുറിച്ചുള്ള വിശദമായ ജീനോമിക് ഡാറ്റ ഞങ്ങൾ ഇവിടെ നൽകുന്നു. ടെർമിനൽ സ്നിപ്പുകൾ ഒഴികെ ഇതിന് ഏതാണ്ട് സമാനമായിരിക്കണം. എന്നാൽ മുകളിൽ പറഞ്ഞ പല കേസുകളിലെയും പോലെ അവ സമാനമാണ്. M223-ലും അതിനു താഴെയുമുള്ള SNP-കൾ ഹൈലൈറ്റ് ചെയ്തിരിക്കുന്നു. കിറ്റ് നമ്പർ. 227059 (ഇംഗ്ലണ്ട്, ശാഖയിലെ ഹാപ്ലോടൈപ്പ് 39): CTS10057+, CTS10058+, CTS10100+, CTS10125+, CTS10362+, CTS109+, CTS11358+, CTS11441+, CTS11441+, CTS2+ CTS16 2+, CTS1977+, CTS1996+, CTS2193+, CTS2392+, CTS2514+, CTS2536+, CTS3296+, CTS3326+, CTS3331+, CTS3431+, CTS3517+, CTS3536+, CTS3654+, CTS3662+, CTS3868+, CTS3996+, CTS4039+, CTS+4088+, CTS4+1TS3, C86, CTS3 8+, CTS444 3+, CTS4740+, CTS4848+, CTS4982+, CTS5286+, CTS5318+, CTS5457+, CTS5532+, CTS5650+, CTS5727+, CTS5908+, CTS6135+, CTS6136+, CTS616+, CTS6265+, CTS6331+, CTS6383+, CTS674+, CTS6800+, CTS6907+, CTS3 CTS1+2, CTS3 CTS20 +, CTS7682+, CTS7831 +, CTS7865+, CTS7922+, CTS7933+, CTS7934+, CTS8243+, CTS8333+, CTS8345+ , CTS8420+, CTS8449+, CTS88+, CTS8876+, CTS8901+, CTS8980+, CTS9183+, CTS9240+, CTS9264+, CTS9266+, CTS+482+, CTS9782+, എഫ്. , F1320+, F1329+, F14 50+, F1460+, F1704+, F1714+, F1753+, F1767+ , F2048+, F2075+, F2142+, F2155+, F2302+, F2345+, F2366+, F2402+, F2587+, F2688+, F2710+, F2794+, F2837+, F2837+, F23913+, F2981+, F23913 35+, F3368+, F3402+, F3556 +, F3692+, F4188+, F719+, F922+, L132+, L15+, L16+, L34+, L35+, L350+, L37+, L403+, L460+, L468+, L470+, L498+, L578+, L59+, L68+, L75++, L75+, L74+, L75+, L72, L75, L72 , L800+, M139+, M168+, M17 0+, M223+, M235+, M294+, M42+, M89+, M94+, P123+, P124+, P126+, P127+, P130+, P135+, P136+, P138+, P14+, P141+, P145+, P145+, P141, P141, P141, P141 P160+, P166+, P187+, P214+ , P215+, P216+, P217+, P219+, P220+, P221+, P222+, P223+, P305+, P95+, PAGES00026+, PAGES00081+, PF1016+, PF+1, PF1029+1, PF1020 061+, PF1092+, PF1097+, PF110+, PF1203+, PF1269+, PF1276+ , PF192+ , PF210+, PF212+, PF223+, PF234+, PF258+, PF2591+, PF2593+, PF2599+, PF2608+, PF2611+, PF2612+, PF2615+, PF2615+ PF2626 F2745+, PF2747+, PF2748+, PF2749+, PF2770+, PF278+, PF292+, PF316+, PF325+ , PF342+ , PF3515+, PF3517+, PF3518+, PF3534+, PF3560+, PF3561+, PF3562+, PF3573+, PF3574+, PF3578+, PF3578+, PF390, P3586 3595+, PF 3596+, PF3600+, PF3604+, PF3605+, PF3607+, PF3611+, PF3612+, PF3616+ , PF3618+, PF3621+ , PF3623+, PF3625+, PF3626+, PF3634+, PF3639+, PF3640+, PF3641+, PF3642+, PF3644+, PF3644+, PF3644+, PF3644+ PF3656, PF3656 F3658+, PF3660+, PF 3664+, PF3666+, PF3667+, PF3669+, PF3671+, PF3672+, PF3675+ , PF3677+, PF3686+, PF3694+ , PF3780+, PF3798+, PF3800+, PF3801+, PF3802+, PF3804+, PF3806+, PF3807+, PF3807+, PF2+81, PF38091, PF38091, PF3809 F3815+, PF3819+, PF3820+, PF 3822+, PF3836+, PF3837+, PF3849+, PF3876+, PF500+ , PF667+, PF719+, PF725+, PF779+ , PF796+, PF803+, PF815+, PF821+, PF840+, PF844+, PF892+, PF937+, PF951+, PF951+, V81950, V,8170 +, V205+, V221+, V241+, V250+, V52 +, V9+, YSC0000056+, YSC0000256+, YSC0000260+, YSC0000265+, YSC0000267+ , YSC0000272+, YSC0000280+, YSC0000281+, YSC0+3, YSC0000029 Z161+, Z162+, Z163+, Z164+, Z165+, Z168+, Z170+, Z172+, Z174+, Z175+, Z176+, Z177+, Z178+, Z179+, Z181+, Z183+, Z186, Z1861, Z1861 0057+, CTS10058+, CTS10100+, CTS10125+ , CTS10834+ , CTS11441+, CTS1977+, CTS2193+, CTS2392+, CTS2514+, CTS3296+, CTS3326+, CTS3517+, CTS3654+, CTS4039+, CTS4039+, CTS4039+, CTS4039+, CTS4048+, CTS4039 CTS4848+, CTS 4982+, CTS5286+, CTS5650+, CTS5727+, CTS5884+, CTS5908+, CTS6136+, CTS616+ , CTS623+, CTS6265+ , CTS6331+, CTS674+, CTS6932+, CTS7329+, CTS7331+, CTS7682+, CTS7831+, CTS7865+, CTS+7934+, C2+CTS34, CTS8333+ 8+, CTS8876+, CTS890 1+, CTS9183+, CTS9240+, CTS9264+, CTS9266+, CTS9482+, CTS9782+ , F1209+, F3692+, F719+ , L132+, L15+, L16+, L181+, L34+, L35+, L350+, L36+, L37+, L403+, L41+, L460+, L460+, L48+, L5, L7, L7, L7, L5 48+, L751+, L755+, L756+ , L758+, L772+, L800+ , M168+, M170+, M213+, M223+, M235+, M258+, M294+, M299+, M42+, M438+, M89+, M94+, P12+, P12+, P12+, P,12+, P,12+, P,12+, P,122+, +, P130+, P133+, P134+ , P135+ , P136+, P138+, P139+ , P14+, P140+, P141+, P143+, P145+, P146+, P148+, P149+, P151+, P157+, P158++, P159+, P160, P160+, P158+, P160+, P160 12+, P214+, P216+, P21 7+ , P218+, P219+, P220+, P221+ , P222+, P223+, P316+, P38+, P95+, PAGES00026+, PAGES00081+, PF2591+, PF2608+, PF2611+, PF2615+, PF2747+, PF2748+, PF2749+, PF2770+, PF3562+, P23+562+, P69 P69 +, PF3641+, PF3644+, PF3651+, PF3652+, PF3657+, PF3658+, PF3660+, PF3664+, PF3666+, PF3667+ , PF3669+, PF3671+, PF3672+, PF3675+, PF3677+, PF3780+, PF3800+, PF3801+, PF3802+, PF3804+, PF+3, PF3809+1, PF3809 815+, PF 3819+, PF3820+, PF3822+, PF3836+, PF3837+, PF3849+, PF3876+, PF6464+, PF6469+, PF6470+ , PF6477+, PF6479+, PF6520+, V241+, V250+, YSC0000207+, YSC0000227+, YSC0000256+, YSC0000270,28 YSC000270,28 Z161+, Z163+, Z164+, Z165+, Z168+, Z170+, Z174+, Z175+, Z176+, Z178+, Z179+, Z184+, Z186+, Z188+, P37-, S2348-, S44422 കിറ്റ് നമ്പർ. 173448 (ഇംഗ്ലണ്ട്, ഹാപ്ലോടൈപ്പ് 44): Z77+, L800+, L801+, V218+, Z186+, Z78+, Z79+, L1198+, Z190+, Z185+, Z171+, Z166+, CTS10057+, CTS10058+, CTS10100+, CTS10125+, CTS10834+, CTS11441+, CTS+2134+, CTS22193+, CTS22193+2 CTS22193+ 26+, CTS3517+, CTS3654+, CTS4039+, CTS4088+, CTS429+, CTS4348+, CTS4437+, CTS4848+ , CTS4982+ , CTS5286+, CTS5650+, CTS5727+, CTS5884+, CTS5908+, CTS6136+, CTS616+, CTS623+, CTS6265+, CTS6331+, CTS6433+, CTS674+, CTS6932+, CTS7329+, CTS7331+, CTS7682+, CTS7831+, CTS7865+, CTS7934+, CTS8333+, CTS8345+, CTS+8420+, CTS1+809, CTS1,8489 3+, CTS9240+, CTS9264+, CTS9266+, CTS9482+, CTS9782+, F1209+, F3692+, F719+ , L104+ , L132+, L15+, L16+, L181+, L34+, L35+, L350+, L36+, L37+, L403+, L41+, L460+, L468+, L498+, L578+, L+8+, L57,451, L59,751 +, L758+, L772+, L800+ , M168+ , M170+, M213+, M223+, M235+, M258+, M294+, M299+, M42+, M438+, M89+, M94+, P108+, P123+, P124+, P125+, P126+, P127+, P129+, P13+ P3, P130+, P16 , P139+, P14+, P140+, P141+ , P143+, P145+, P146+, P148+, P149+, P151+, P157+, P158+, P159+, P160+, P161+, P163+, P166+, P187+, P212+, P218, P218, P219, 20+, P221+, P222+, P223+ , P305+ , P316+, PAGES00026+, PAGES00081+, PF2591+, PF2608+, PF2611+, PF2615+, PF2624+, PF2747+, PF2748+, PF2748+, PF2748+, PF27273, PF272749 F3626+, PF3639+, PF3640+, PF3641+, PF3644+, PF3651+, PF3652+, PF3657+, PF3658+, PF3660+ , PF3664+ , PF3666+, PF3667+, PF3669+, PF3671+, PF3672+, PF3675+, PF3677+, PF3780+, PF3800+, PF3801+, PF3801+, PF3801+, PF3902, PF3802, PF3802 F3812+, PF 3814+, PF3815+, PF3819+, PF3820+, PF3822+, PF3836+, PF3837+, PF3849+ , PF3876+, PF6464+ , PF6469+, PF6470+, PF6477+, PF6479+, PF6520+, PK1+, SRY10831+, V221+, V241+, V250+2 YSC0+2 YSC0+2 YSC0+2 YSC0+2 YS00, YS002, YSC02, YSC002 , YSC0000272+, Z163+, Z164+, Z165+, Z168+, Z170+, Z174+, Z175+, Z178+, Z179+ , Z184+, Z185+, Z188+, Z190+, Z79+, L707+, M223+, M253-, M26-, M284-, M379-, P78-, P95-, S2348-, S4442- കിറ്റ് നമ്പർ. 18187 (പോർച്ചുഗൽ, ഹാപ്ലോടൈപ്പ് 45): CTS674+, CTS6800+, CTS6907+, CTS6932+, CTS7329+, CTS7331+, CTS7502+, CTS7682+, CTS7831+, CTS3+ CTS7865, CTS3+ CTS7865 TS 8243+, CTS8333+, CTS8345+, CTS8420+, CTS8449+, CTS88+, CTS8876+, CTS8901+, CTS8980+, CTS9183+, CTS9240+, CTS9264+, CTS9266+, CTS9482+, CTS9782+, CTS9828+, F1046+, F1209+, F1302+, F1342+, F13420+, F13420+, F13410 04+, F171 4+, F1753+, F1767+, F2048+, F2075+, F2142+, F2155+, F2302+, F2345+, F2366+, F2402+, F2587+, F2688+, F2710+, F2794+, F2837+, F2985+, F2993+, F3111+, F3136+, F3336+, F3336+, F3336+, F333562, F33356 +, F4188+, F719+ , F922+, L132+, L15+, L16+, L34+, L35+, L350+, L37+, L403+, L41+, L460+, L468+, L470+, L498+, L578+, L59+, L68+, L748+, L751+, L755+, L756+, L73+ M, L756+, L73+ M1 0+, M223+, M235+, M294+, M42+, M89+, M94+, P123+, P124+, P126+, P127+, P130+, P135+, P136+, P138+, P14+, P141+, P145+, P145+, P141, P141, P141, P141 P160+, P166+, P187+, P214+ , P215+, P216+, P217+, P219+, P220+, P221+, P222+, P223+, PF1016+, PF1029+, PF1031+, PF1040+, PF1046+, PF+10161, P+10161 203+, PF1269+, PF1276+, PF192+, PF210+, PF212+, PF223+ , PF234+ , PF258+, PF2591+, PF2593+, PF2599+, PF2608+, PF2611+, PF2615+, PF2624+, PF263+, PF2643+, PF272+, PF272+ 9+, PF277 0+, PF278+, PF292+, PF316+, PF325+, PF342+, PF3515+, PF3517+ , PF3518+, PF3534+ , PF3560+, PF3561+, PF3562+, PF3573+, PF3574+, PF3578+, PF3586+, PF3588+, PF3590+, PF3590+, PF3590+, PF3590+, PF60, P3594 F3604+, PF3605+, PF 3607+, PF3611+, PF3612+, PF3616+, PF3618+, PF3621+, PF3623+ , PF3625+, PF3626+, PF3634+ , PF3639+, PF3640+, PF3641+, PF3642+, PF3644+, PF3651+, PF3652+, PF3654+, PF3654+, PF3654+, PF3654+ F3666+, PF3667+, PF3669+, PF 3671+, PF3672+, PF3675+, PF3677+, PF3686+, PF3694+ , PF3780+, PF3798+, PF3800+, PF3801+ , PF3802+, PF3804+, PF3806+, PF3807+, PF3809+, PF3811+, PF3812+, PF3812+, PF2+381, PF3814, PF3814 F3822+, PF3836+, PF3837+, PF3849+, PF 3876+, PF500+, PF667+, PF719+, PF725+ , PF779+, PF796+, PF803+, PF815+, PF821+ , PF840+, PF844+, PF892+, PF937+, PF951+, PF954+, PF970+, V186++, V186++, V180 V180, V180, V189 , YSC0000256+, YSC0000260+, YSC0000265+, YSC 0000267+, YSC0000272+, YSC0000280+ , YSC0000281+, YSC0000298+, YSC0000300+, Z161+, Z162+, Z163+, Z164+, Z165+, M170+, M223+, M258+, P38+, L801+, Z186+, Z76+, L1316+, CTS10057+, CTS10058+, CTS10+58+, CTS2+1010, CTS1010, CTS1010, , CTS11358+, CTS11441+, CTS11575+, CTS11726+, CTS125+, CTS12632+, CTS1996+ , CTS2193+, CTS2392+, CTS2514+, CTS2536+, CTS3296+, CTS3326+, CTS3331+, CTS3431+, CTS3517+, CTS3536+, CTS3654+, CTS++ CTS38, CTS3662 4088+, CTS429+, CTS4314+, CTS4348+, CTS4364+, CTS4368+, CTS4443+, CTS4740+, CTS4848+ , CTS4982+ , CTS5286+, CTS5318+, CTS5457+, CTS5532+, CTS5650+, CTS5727+, CTS5908+, CTS6135+, CTS6136+, CTS616+, CTS3+ CTS363+ 8+ CTS6265 CTS6433+, F3692+, F719+, L104+, L132+, L15+, L16+, L181+, L34+, L35+, L350+, L36+, L37+, L403+, L41+, L460+, L468+, L+8+5, L597,5 +, L756+, L758+, L772+ , L800+, M168+, M170+, M213+, M223+, M235+, M258+, M294+, M299+, M42+, M438+, M89+, M94+, P123+, P124+, P126+, P126+, P12, P12 P133+, P134+, P135+, P136+, P138+ , P139+, P14+, P140+, P141+, P145+, P146+, P148+, P149+, P151+, P157+, P158+, P159+, P160+, P161+, P16+ P163, P163, P163, P12 +, P21 7+, P218+, P219+, P220+, P221+ , P222+, P223+, P316+, PAGES00026+, PAGES00081+, PF2591+, PF2608+, PF2611+, PF2615+, PF2747+, PF2747+, PF272749, PF2727 3833+, Z2198 +, CTS8444+, CTS6497+, CTS10057+, CTS10058+, CTS10100+, CTS10125+, CTS10834+ , CTS11441+, CTS2134+ , CTS2193+, CTS2392+, CTS2514+, CTS3296+, CTS3326+, CTS3517+, CTS3654+, CTS4039+, CTS4088+, CTS4088+, CTS74348+, CTS429 TS4982+, CTS5286+, CTS5 650+, CTS5727+, CTS5884+, CTS5908+, CTS6136+, CTS616+, CTS623+ , CTS6265+, CTS6331+, CTS674+ , CTS6932+, CTS7329+, CTS7331+, CTS7682+, CTS7831+, CTS7934+, CTS8333+, CTS8345+, CTS+CTS8489, CTS8420 01+, CTS9183+, CTS9240+, CTS92 64+, CTS9266+, CTS9482+, CTS9782+, PF3562+, PF3574+ , PF3626+, PF3639+, PF3640+, PF3641+ , PF3644+, PF3651+, PF3652+, PF3657+, PF3658+, PF3660+, PF3664+, PF3664+, PF3664+, PF3666+ PF3666, PF3666 F3672+, PF3675+, PF3677+, PF3780+, PF 3800+, PF3801+, PF3802+, PF3804+, PF3809+ , PF3811+, PF3812+, PF3814+, PF3815+, PF3819+ , PF3820+, PF3822+, PF3836+, PF3837+, PF3849+, PF3876+, Z++ Z+176+, Z1174+, Z1175,176 8+, Z179+, P19+, Z181+, Z183+, Z184+, Z186+, V22 1+ , V241+, V250+, YSC0000207+, YSC0000227+, YSC0000256+, YSC0000272+ , Z163+, Z164+, Z165+, Z168+, Z170+, Z175+ P, Z174+, Z174+, Z4 469+, PF6470+, PF6477+, PF6479+, PF6520+, Z168+, Z170+, Z188+, Z7 7+, Z188+, F1209+, F3406-, M161-, Z190 -, Z63-, Z79-, L1317-, Z171-, S2348-, S4442-, Z185-, Z187-, L1201-, P70,18 -, PF4225-, DF5-, PF3292-, CTS661-, L1198-, Z190-, Z2059-, Z2084-, Z2074-, L1272-, ZS6-, ZS7-, ZS11-, ZS14-, ZS15-, ZS16-, ZS18-, ZS26-, L1290-, Z78-, Z79-, M284-, M379-, P78-, P95-, L380-, M26-, M307-, M72-, M227-, M253-, M21-, Z166- കിറ്റ് നമ്പർ. B3411 (സ്വിറ്റ്സർലൻഡ്, ഹാപ്ലോടൈപ്പ് 46): L801+, CTS6433+, M223+, L1201-, Z190-, Z78-, CTS5332- കിറ്റ് നമ്പർ. 23157 (സ്കോട്ട്ലൻഡ്, ഹാപ്ലോടൈപ്പ് 51): P19+, M170+, M258+, M223+, P19+, P38+, M284-, M379-, P78-, P95-, P30-, P37-, M227-, M253-, M26-, M307-, M72-, M21-, M161- കിറ്റ് നമ്പർ. 174472 (സ്വീഡൻ, ഹാപ്ലോടൈപ്പ് 52): M284-, L126-, M379-, P78-, P95- ഹാപ്ലോഗ് ഗ്രൂപ്പ് I2c KLIN ID00005 FTDNA-യിലെ ആഴമേറിയ ഉപവിഭാഗമായ I2c-L596 സബ്ക്ലേഡ് തിരിച്ചറിഞ്ഞിട്ടില്ലെന്ന് റിപ്പോർട്ടുണ്ട്. 14 24 15 10 12 13 11 13 12 12 11 28 – 18 8 8 11 11 24 15 21 31 11 11 15 15 – 10 10 19 21 15 13 17 17 34 34 14 10 12 8 16 16 8 13 10 8 10 9 12 21 21 16 11 12 13 13 8 14 25 21 12 13 11 13 11 12 12 11 I2c-L596-നേക്കാൾ ആഴത്തിൽ സബ്ക്ലേഡ് നിർണ്ണയിച്ചിട്ടില്ലാത്തതിനാൽ, ഹാപ്ലോഗ് ഗ്രൂപ്പ് I-ൽ യഥാർത്ഥത്തിൽ നിർവചിച്ചിരിക്കുന്ന സബ്ക്ലേഡുകളുടെ ശൃംഖല വളരെ ചെറുതാണ്: I-M170 > I2-M438 > I2c-L596. 67-മാർക്കർ ഹാപ്ലോടൈപ്പുകളുടെ മരത്തിൽ അവതരിപ്പിച്ച ഹാപ്ലോടൈപ്പിൻ്റെ സ്ഥാനം കാണിക്കുന്നത് പോലെ, FTDNA-യിലെ തിരിച്ചറിയൽ ശരിയായി നടപ്പിലാക്കി. എന്നാൽ ഇനിപ്പറയുന്ന അവരോഹണ ഉപവിഭാഗങ്ങൾ സാധ്യമാണ്, അവ ഇതുവരെ നിർണ്ണയിച്ചിട്ടില്ല:
ഇപ്പോൾ 9 ഹാപ്ലോടൈപ്പുകളുടെ ഒരു ശാഖയെക്കുറിച്ച് (മൂന്ന് ജോർജിയയിൽ നിന്ന് [427, 451, 452 എണ്ണം], ബെലാറസിൽ നിന്ന് രണ്ട്, ലിത്വാനിയ, ഓസ്ട്രിയ, തുർക്കി എന്നിവിടങ്ങളിൽ നിന്ന് ഓരോന്നും). ഈ ശാഖയുടെ അടിസ്ഥാന ഹാപ്ലോടൈപ്പ് (അവതരിപ്പിച്ച ഹാപ്ലോടൈപ്പിൽ നിന്നുള്ള മ്യൂട്ടേഷൻ വ്യത്യാസങ്ങൾ അടയാളപ്പെടുത്തിയിരിക്കുന്നു) 14 24 15 10 12 13 11 13 11 13 11 29 – 18 8 8 11 11 24 15 20 32 11 14 15 16 – 11 10 19 21 15 13 17 19 34 36 12 10 12 8 16 16 8 12 10 8 10 9 12 21 21 16 11 12 13 14 8 13 25 20 12 13 11 13 11 12 12 11 ശാഖയുടെ എല്ലാ 9 ഹാപ്ലോടൈപ്പുകളിലും ഈ അടിസ്ഥാന ഹാപ്ലോടൈപ്പിൽ നിന്ന് 123 മ്യൂട്ടേഷനുകൾ ഉണ്ട്, അതായത് ശാഖയുടെ പൊതു പൂർവ്വികൻ 123/9/0.12 = 114 → 129 പരമ്പരാഗത തലമുറകൾക്ക് മുമ്പ് അല്ലെങ്കിൽ 3225 ± 430 വർഷം മുമ്പ് ജീവിച്ചിരുന്നു എന്നാണ്. അവതരിപ്പിച്ച ഹാപ്ലോടൈപ്പ് അടിസ്ഥാനം ഒന്നിൽ നിന്ന് 17 മ്യൂട്ടേഷനുകൾ വ്യത്യാസപ്പെട്ടിരിക്കുന്നു (പ്രത്യേക നിയമങ്ങൾക്കനുസൃതമായി നിരവധി മ്യൂട്ടേഷനുകൾ കണക്കാക്കുന്നു), ഇത് കണക്കുകൂട്ടൽ പിശകിനുള്ളിൽ സാധാരണ പൂർവ്വികനിൽ നിന്ന് ഏകദേശം ഒരേ സമയം നൽകുന്നു. ബിസി രണ്ടാം സഹസ്രാബ്ദത്തിൻ്റെ അവസാനത്തിൽ ജീവിച്ചിരുന്ന ഹാപ്ലോഗ് ഗ്രൂപ്പ് I2c യുടെ ശാഖയുടെ പൊതു പൂർവ്വികൻ്റെ നേരിട്ടുള്ള പിൻഗാമിയാണ് ID00005 എന്ന് ഇത് കാണിക്കുന്നു.
L596 സബ്ക്ലേഡിൻ്റെയും അതിൻ്റെ താഴത്തെ സബ്ക്ലേഡുകളുടെയും ചരിത്രം എത്ര സങ്കീർണ്ണവും ദാരുണവുമാണെന്ന് മനസിലാക്കാൻ, L596 സബ്ക്ലേഡ് തന്നെ ഏകദേശം 21,300 വർഷങ്ങൾക്ക് മുമ്പ് (Y ക്രോമസോമിൽ കാണിക്കുന്ന SNP-കളുടെ ഗോവണി പോലെ) ഉടലെടുത്തതായി ഞങ്ങൾ ശ്രദ്ധിക്കുന്നു, പക്ഷേ അത് പൂർണ്ണമായും നശിച്ചു. യൂറോപ്പിൻ്റെ ചരിത്രത്തിൽ, L596-ൻ്റെ അതിജീവിച്ച ഏതാനും വാഹകരിൽ നിന്ന് വീണ്ടും പുനർജനിച്ചു. തൽഫലമായി, ID00005 എന്ന ഉപശാഖയുടെ പൂർവ്വികൻ ഏകദേശം 3225 വർഷം മുമ്പ് ജീവിച്ചിരുന്നു, മുകളിലുള്ള ഡയഗ്രാമിലെ അയൽ ഉപശാഖ (ഏഴ് ഹാപ്ലോടൈപ്പുകളുടെ) - ഏകദേശം 2475 വർഷങ്ങൾക്ക് മുമ്പ്, മുകളിലെ ഉപശാഖ (ഒമ്പത് ഹാപ്ലോടൈപ്പുകൾ) - 2800 വർഷം മുമ്പ്, കൂടാതെ എല്ലാ ശാഖകളും. ഒരുമിച്ച് - 5950 ± 645 വർഷം മുമ്പ്. ജീനോമിക് വിശകലനം കാണിക്കുന്നത് താഴത്തെ ഉപക്ലേഡ് I2c1 15,500 വർഷങ്ങൾക്ക് മുമ്പ് രൂപപ്പെട്ടതാണെന്ന്; താഴെയുള്ള അടുത്ത ഉപവിഭാഗം, I2c1a-L1251 - 10,100 വർഷങ്ങൾക്ക് മുമ്പ്, ബാക്കിയുള്ളവയെക്കുറിച്ച് ഇതുവരെ വിവരങ്ങളൊന്നുമില്ല. L596 ൻ്റെ മൂന്ന് ശാഖകൾ ഇതുവരെ കണ്ടെത്തിയിട്ടുണ്ട്, ഓരോന്നും ഏകദേശം 3200-2500 വർഷങ്ങൾക്ക് മുമ്പുള്ളതാണ്.
അതിനാൽ, ഇന്നത്തെ നമ്മുടെ അറിവിൻ്റെ അവസ്ഥ സംഗ്രഹിക്കാം. ഹാപ്ലോഗ് ഗ്രൂപ്പ് I വളരെ പുരാതനമാണ്, 40 ആയിരത്തിലധികം വർഷങ്ങൾക്ക് മുമ്പ് രൂപീകരിച്ചു, അവിടെ ഇപ്പോഴും അജ്ഞാതമാണ്, പക്ഷേ മിക്കവാറും യൂറോപ്പിൽ. ഏകദേശം 30 ആയിരം വർഷങ്ങൾക്ക് മുമ്പ് ഇത് I1, I2 എന്നീ ഉപക്ലേഡുകളായി വ്യതിചലിച്ചു, രണ്ടാമത്തേത് ഏകദേശം 21,300 വർഷങ്ങൾക്ക് മുമ്പ് I2a, I2b, I2c-L596 എന്നീ ഉപക്ലേഡുകളായി വ്യതിചലിച്ചു. ഹാപ്ലോഗ് ഗ്രൂപ്പുകൾ I, I2 എന്നിവ മധ്യ യൂറോപ്പിലെയും സ്വീഡനിലെയും 7 ആയിരം വർഷങ്ങൾക്ക് മുമ്പുള്ള പുരാതന ഫോസിൽ അസ്ഥി അവശിഷ്ടങ്ങളിൽ കണ്ടെത്തി. കുറച്ച് കഴിഞ്ഞ്, മൂന്ന് ശാഖകളുടെ ഒരു പൊതു പൂർവ്വികൻ ജീവിച്ചിരുന്നു, I2c-L596, അവരുടെ പിൻഗാമികൾ നിലവിൽ ബ്രിട്ടീഷ് ദ്വീപുകൾ മുതൽ തുർക്കി വരെ താമസിക്കുന്നു. എന്നാൽ ഏകദേശം 4500 വർഷങ്ങൾക്ക് മുമ്പ്, യൂറോപ്പിലെ എല്ലാവരേയും പോലെ ഹാപ്ലോഗ് ഗ്രൂപ്പ് I ൻ്റെ വാഹകരുടെ ജീവിതം കുത്തനെ തടസ്സപ്പെട്ടു, മിക്കവാറും എല്ലാ ഹാപ്ലോഗ് ഗ്രൂപ്പുകളും (R1b ഒഴികെ) യൂറോപ്പിൽ നിന്ന് അപ്രത്യക്ഷമാവുകയോ യൂറോപ്പിൻ്റെ പെരിഫറൽ പ്രദേശങ്ങളിലേക്ക് തിരികെ പോകുകയോ ചെയ്തു. പശ്ചിമേഷ്യയും കോക്കസസും. ഈ ഹാപ്ലോഗ് ഗ്രൂപ്പുകൾ, വ്യക്തിഗത പ്രതിനിധികൾ എന്ന നിലയിൽ, അതിജീവനത്തിൻ്റെ ദീർഘവും വേദനാജനകവുമായ ഒരു കാലഘട്ടത്തിലൂടെ കടന്നുപോകുന്നു, അതിജീവിച്ചവർ പെരുകാൻ തുടങ്ങുന്നു, ഇത് പുതിയ ശാഖകളുടെ പൊതു പൂർവ്വികരെ രൂപപ്പെടുത്തുന്നു. 3200-2500 വർഷങ്ങൾക്ക് മുമ്പ് “വളരാൻ” തുടങ്ങിയ I2c-L596 സബ്ക്ലേഡിൻ്റെ വാഹകരും അവരിൽ ഉൾപ്പെടുന്നു, അതായത്, ബിസി ഒന്നാം സഹസ്രാബ്ദത്തിൻ്റെ മധ്യത്തിൽ. ഏതാണ്ട് അതേ സമയം, ഹാപ്ലോഗ് ഗ്രൂപ്പ് I2a പുനരുജ്ജീവിപ്പിക്കാൻ തുടങ്ങി, അതിൻ്റെ പൊതു പൂർവ്വികൻ 2300 വർഷങ്ങൾക്ക് മുമ്പ് കിഴക്കൻ യൂറോപ്പിലും ഏകദേശം 5000 വർഷങ്ങൾക്ക് മുമ്പ് ബ്രിട്ടീഷ് ദ്വീപുകളിലും ജീവിച്ചിരുന്നു. കുറച്ച് മുമ്പ്, ഹാപ്ലോഗ് ഗ്രൂപ്പ് I1 പുനരുജ്ജീവിപ്പിക്കാൻ തുടങ്ങി, ഏകദേശം 3,700 വർഷങ്ങൾക്ക് മുമ്പ് ജീവിച്ചിരുന്ന ഒരു പൊതു പൂർവ്വികൻ. ഇക്കാര്യത്തിൽ, ഈ ഹാപ്ലോഗ് ഗ്രൂപ്പിൻ്റെ ഇന്നത്തെ വാഹകരുമായി ബന്ധപ്പെട്ട് I2c-L596 സബ്ക്ലേഡിന് ഏറ്റവും പുരാതനമായ വേരുകളുണ്ട് - ഏകദേശം 6000 വർഷങ്ങൾക്ക് മുമ്പ്. L596 ൻ്റെ താഴത്തെ ഉപവിഭാഗങ്ങളെ ഹാപ്ലോടൈപ്പുകളുടെ അനുബന്ധ ശാഖകളായി തിരിച്ചിരിക്കുന്നു എന്നത് സന്തോഷകരമാണ് (മുകളിലുള്ള ഡയഗ്രം കാണുക). ഇതിനർത്ഥം, മോസ്കോ ലബോറട്ടറി ഓഫ് ഡിഎൻഎ വംശാവലിയിൽ ഈ താഴ്ന്ന സബ്ക്ലേഡുകൾ മതിയായ ആളുകളിൽ (ഉദാഹരണത്തിന്, അമ്പത്) തിരിച്ചറിയുമ്പോൾ, ഉപക്ലേഡുകളിലേക്ക് ഹാപ്ലോടൈപ്പുകളുടെ കൂടുതൽ വിശദമായ അസൈൻമെൻ്റ് നടത്താനും കൂടുതൽ വിശദമായി മനസ്സിലാക്കാനും ഞങ്ങൾക്ക് കഴിയും. ഉപക്ലേഡുകളുടെ പ്രധാന ഭൂമിശാസ്ത്രം. വ്യക്തിഗത mtDNA വ്യാഖ്യാന പരിശോധന സ്ത്രീ ഹാപ്ലോഗ് ഗ്രൂപ്പുകളിലേക്ക് വ്യക്തിഗത വ്യാഖ്യാനങ്ങളുടെ പരിധി വിപുലീകരിക്കുന്നത് തീർച്ചയായും ഉപയോഗപ്രദമാകും, കൂടാതെ സ്ത്രീകളിൽ നിന്നും പുരുഷന്മാരിൽ നിന്നുമുള്ള mDNA അവ പരിശോധിക്കാൻ വിശകലനം ചെയ്യാവുന്നതാണ്. ഒരു മനുഷ്യൻ തൻ്റെ അമ്മയിൽ നിന്ന് രക്തത്തിൽ mtDNA സ്വീകരിക്കുന്നു, പക്ഷേ mtDNA കൂടുതൽ കൈമാറുന്നില്ല എന്നതാണ് വസ്തുത, കാരണം ബീജസങ്കലനത്തിനു ശേഷം (അല്ലെങ്കിൽ അതിനിടയിൽ) ബീജത്തിൻ്റെ മൈറ്റോകോണ്ട്രിയ നശിപ്പിക്കപ്പെടുന്നു. ബീജത്തിൽ ഏകദേശം ഒരു ഡസനോളം മൈറ്റോകോൺഡ്രിയ അടങ്ങിയിരിക്കുന്നു, കാരണം വാലിൻ്റെ ഭ്രമണത്തിന് ഊർജ്ജം ആവശ്യമാണ്, അത് നൽകുന്നത് മൈറ്റോകോൺഡ്രിയയാണ്. ബീജം അതിൻ്റെ ലക്ഷ്യത്തിലെത്തുമ്പോൾ, മൈറ്റോകോണ്ട്രിയ ഉപേക്ഷിക്കപ്പെടുന്നു. നിങ്ങൾ പുരുഷന്മാരെ തിരിച്ചറിയുന്നുണ്ടോ? വളരെ അപൂർവ സന്ദർഭങ്ങളിൽ, ഒരു മൈറ്റോകോൺഡ്രിയ ഇപ്പോഴും കടന്നുപോകുന്നു, പക്ഷേ ഇത് പ്രതീക്ഷിക്കുന്ന അമ്മയുടെ ശരീരവുമായി ബന്ധപ്പെട്ട് ഒരു വിദേശ മൈറ്റോകോൺഡ്രിയയാണ്, ഇത് ഒരു ചട്ടം പോലെ, നന്നായി അവസാനിക്കുന്നില്ല. ഇത് പലപ്പോഴും അക്യൂട്ട് പാത്തോളജികളിൽ അവസാനിക്കുന്നു. ചുരുക്കത്തിൽ, mtDNA അമ്മയിൽ നിന്ന് പെൺമക്കളിലേക്ക് തലമുറകളുടെ മുഴുവൻ ശൃംഖലയിലൂടെയും, പതിനായിരക്കണക്കിന്, ലക്ഷക്കണക്കിന് വർഷങ്ങൾ, ദശലക്ഷക്കണക്കിന് വർഷങ്ങൾ എന്നിങ്ങനെ കൈമാറ്റം ചെയ്യപ്പെടുന്നു. ആൺമക്കൾക്കും - ഒരു തലമുറയ്ക്ക് മാത്രം. ആൺമക്കൾ Y ക്രോമസോം അവരുടെ ആൺമക്കൾക്ക് കൈമാറുന്നു, mtDNA ആർക്കും കൈമാറരുത് - ആൺമക്കളോ പെൺമക്കളോ അല്ല, എന്നാൽ അവരുടെ മറ്റ് 22 ക്രോമസോമുകൾ, പുനഃസംയോജനം, എല്ലാവർക്കും കൈമാറുന്നു. അതിനാൽ, ഞാൻ ആവർത്തിക്കുന്നു, സ്ത്രീകളുടെയും പുരുഷന്മാരുടെയും ഡിഎൻഎയിൽ mtDNA ടെസ്റ്റുകൾ നടത്താം. രണ്ട് സാഹചര്യങ്ങളിലും, ഫലം ഒന്നുതന്നെയാണ് - അമ്മയുടെ mtDNA യുടെയും മുമ്പത്തെ മുഴുവൻ mtDNA ശൃംഖലയുടെയും എല്ലാ കുമിഞ്ഞുകൂടിയ മ്യൂട്ടേഷനുകളും തിരിച്ചറിയൽ. ഇത് നൽകുന്ന കാര്യങ്ങൾ പല പ്രസിദ്ധീകരണങ്ങളിലും, പ്രത്യേകിച്ചും, ഞങ്ങൾക്ക് ഏറ്റവും അനുയോജ്യമായ രൂപത്തിൽ - “ഇൻ്റർനെറ്റ്” എന്ന പുസ്തകങ്ങളിൽ വിവരിച്ചിരിക്കുന്നു. ഒരു ഗവേഷകനിൽ നിന്നുള്ള കുറിപ്പുകൾ" (2010), പേജ്. 368-379, അല്പം വിപുലീകരിച്ച രൂപത്തിൽ - "സ്ലാവുകളുടെ ഉത്ഭവം" (2013), പേജ് 265-284. mtDNA-യുടെ വ്യക്തിഗത വ്യാഖ്യാനത്തിനായുള്ള കാമ്പെയ്ൻ ആരംഭിക്കുന്നതിന് മുമ്പ്, ഞങ്ങളുടെ സഹപ്രവർത്തകരിലൊരാൾ നിർമ്മിച്ച ഒരു ട്രയൽ പതിപ്പ് ഞങ്ങൾ വാഗ്ദാനം ചെയ്യുന്നു. ഇതൊരു പരീക്ഷണ പതിപ്പായതിനാൽ, വ്യാഖ്യാനത്തിൻ്റെ രചയിതാവിൻ്റെ പേര് ഇപ്പോൾ മറച്ചിരിക്കുന്നു. ഈ ഘട്ടത്തിൽ, ഉൽപ്പന്നം കുറഞ്ഞത് സ്വീകാര്യമായി കണക്കാക്കേണ്ടത് പ്രധാനമാണ്, അതിലും മികച്ചത്, നല്ലതോ മികച്ചതോ ആണ്. അതിനാൽ ഇത് സാധ്യതയുള്ള ഉപഭോക്താക്കളുടെയും എല്ലാ വായനക്കാരുടെയും പരിഗണനയ്ക്കാണ്. അതിനാൽ, ഒരു ടെസ്റ്റ് ഓപ്ഷനായി, ഇനിപ്പറയുന്നവ പരിഗണിക്കുക:
അവതരിപ്പിച്ച ഡാറ്റ, പാലിയോലിത്തിക്ക് (ശിലായുഗം) മുതൽ ബിസി ആയിരക്കണക്കിന് വർഷത്തെ ചക്രവാളം വരെ മാത്രം ആഴത്തിൽ ജനുസ്സിൻ്റെ ചരിത്രം കണ്ടെത്താൻ ഞങ്ങളെ അനുവദിക്കുന്നു. ഒന്നാമതായി, സ്ത്രീ മൈറ്റോകോൺഡ്രിയൽ ഡിഎൻഎയ്ക്ക് ഉയർന്ന വേരിയബിൾ പ്രദേശങ്ങളുടെ നീളം വളരെ കുറവാണ്, അതിനാൽ ഇത് വളരെ കുറച്ച് വിവരദായകമാണ് എന്നതാണ് ഇതിന് കാരണം. രണ്ടാമതായി, അവതരിപ്പിച്ച ഡാറ്റയിൽ ലഭ്യമായ വിശദാംശങ്ങൾ അടങ്ങിയിട്ടില്ല - ഉദാഹരണത്തിന്, H1c സബ്ക്ലേഡ്, ചുവടെ കാണിച്ചിരിക്കുന്നതുപോലെ, അവതരിപ്പിച്ച മ്യൂട്ടേഷൻ ചിത്രം ഉൾപ്പെടുന്നതാണ്, 20-ലധികം താഴ്ന്ന സബ്ക്ലേഡുകൾ അടങ്ങിയിരിക്കുന്നു. അതിനാൽ, Y ക്രോമസോമിലെ ഡാറ്റയുടെ വ്യാഖ്യാനവുമായി താരതമ്യപ്പെടുത്തുമ്പോൾ, ജനുസ്സിൻ്റെ ചരിത്രം കൂടുതൽ ഏകദേശ കണക്കിൽ മാത്രമേ പുനർനിർമ്മിക്കാൻ കഴിയൂ. RSRS-ഉം HVR1 ഉം HVR2 ഉം തമ്മിലുള്ള വ്യത്യാസങ്ങളുടെ വിശകലനം കാണിക്കുന്നത്, അവതരിപ്പിച്ച ഹാപ്ലോടൈപ്പ് മൈറ്റോകോൺഡ്രിയൽ ഹാപ്ലോഗ് ഗ്രൂപ്പ് H1c യുടെതാണെന്ന് കാണിക്കുന്നു. മുകളിൽ സൂചിപ്പിച്ച കാരണങ്ങളാൽ, നൽകിയിരിക്കുന്ന സാമ്പിൾ മറ്റ് പല mtDNA-കളുടെ ഡാറ്റയുമായി പൊരുത്തപ്പെടുന്നു: familytreedna.com ഡാറ്റാബേസിൽ HVR1, HVR2 എന്നിവയ്ക്ക് സമാനമായ RSRS മൂല്യങ്ങളുള്ള ഹാപ്ലോടൈപ്പുകളുടെ വാഹകരുണ്ട്, അവയിൽ ഭൂരിഭാഗവും വടക്കും കിഴക്കും ഉള്ളവയാണ്. യൂറോപ്പ്, ഉദാഹരണത്തിന്, അനെല സരോക്കയ്ക്ക് (ലിത്വാനിയ) mtDNA. ഹാപ്ലോടൈപ്പ് ചരിത്രം. സ്ത്രീ ഹാപ്ലോഗ് ഗ്രൂപ്പ് എച്ച് ഏകദേശം 30 ആയിരം വർഷങ്ങൾക്ക് മുമ്പ് പടിഞ്ഞാറൻ ഏഷ്യയിൽ നിന്ന് ഉത്ഭവിച്ചതായി വിശ്വസിക്കപ്പെടുന്നു, ഏകദേശം 20-25 ആയിരം വർഷങ്ങൾക്ക് മുമ്പ് യൂറോപ്പിൽ എത്തി, ഭൂഖണ്ഡത്തിൻ്റെ തെക്കുപടിഞ്ഞാറായി ഫ്രാങ്കോ-കാൻ്റാബ്രിയൻ മേഖലയിലേക്ക് വ്യാപിച്ചു, ഇത് മിക്കവാറും മഡലീൻ സംസ്കാരവുമായി യോജിക്കുന്നു. . 20-13 ആയിരം വർഷങ്ങൾക്ക് മുമ്പുള്ള അവസാന ഗ്ലേഷ്യൽ കാലഘട്ടത്തിൽ, വടക്കൻ, മധ്യ യൂറോപ്പിലെ ഭൂരിഭാഗം പാലിയോലിത്തിക് വാസസ്ഥലങ്ങളും നശിച്ചു, അതിനാൽ ഹാപ്ലോഗ് ഗ്രൂപ്പ് എച്ച് പ്രതിനിധികൾ സ്പെയിനിൻ്റെ വടക്ക് ഭാഗത്ത് മാത്രമേ അതിജീവിച്ചുള്ളൂ (അതിനാൽ, നിലവിൽ ഈ ഹാപ്ലോഗ് ഗ്രൂപ്പ് ഉയർന്ന ആവൃത്തി, 50% ൽ കൂടുതൽ, പ്രാദേശിക ഓട്ടോക്ത്തോണസ് ജനസംഖ്യയുടെ ചൈന-കൊക്കേഷ്യൻ അന്യഗ്രഹജീവികളുടെ സ്വാംശീകരണ സമയത്ത് ഈ പ്രദേശത്ത് രൂപംകൊണ്ട ബാസ്ക്കുകൾക്കിടയിൽ കണ്ടെത്തി). ഈ (ഫോസിൽ) ഹാപ്ലോഗ് ഗ്രൂപ്പിനെ ലാ പാസീഗ, ലാ ചോറ (മഡലീൻ) ഗുഹകളിൽ തിരിച്ചറിഞ്ഞു. പിന്നീടുള്ള ചക്രവാളങ്ങളിൽ, മെസോ-നിയോലിത്തിക്ക് മുതൽ വടക്കൻ യൂറോപ്പിലും റഷ്യൻ സമതലത്തിലും - സെർട്ടിയയിൽ (സ്മോലെൻസ്ക് മേഖല) ആരംഭിച്ച് ഹാപ്ലോഗ് ഗ്രൂപ്പിൻ്റെ ആദ്യകാല സാന്നിധ്യം ശ്രദ്ധിക്കേണ്ടതാണ്, അവിടെ സെർട്ടിയ VIII ലെയറിൽ ഹാപ്ലോടൈപ്പ് H ൻ്റെ ഒരു കാരിയർ കണ്ടെത്തി. പുരുഷ ഹാപ്ലോടൈപ്പ് R1a1 (IV മില്ലേനിയം BC) യുമായി ചേർന്ന്. ഈ പുരാവസ്തു സംസ്കാരത്തെ ഫണൽ ബീക്കർ സംസ്കാരത്തിൻ്റെ വിദൂരപരിധിയായി കണക്കാക്കാം. കൂടാതെ, ബിസി 5-4 സഹസ്രാബ്ദങ്ങളിലെ ശ്മശാനങ്ങളിലും സമാനമായ ഹാപ്ലോടൈപ്പുകൾ കണ്ടെത്തി. ഡൊനെറ്റ്സ്ക് മേഖലയിലും യുഷ്നി ഒലെനി ഓസ്ട്രോവ് ശ്മശാനത്തിലും (പുരുഷ ഹാപ്ലോഗ് ഗ്രൂപ്പായ R1a1* തിരിച്ചറിയപ്പെട്ടിരുന്നു). സബ്ക്ലേഡ് H1c. ഹാപ്ലോടൈപ്പ് H1c സബ്ക്ലേഡിൻ്റേതാണോ എന്ന് നിർണ്ണയിക്കാൻ T477C സൂചകം ഞങ്ങളെ അനുവദിക്കുന്നു. ഏകദേശം 13 ആയിരം വർഷങ്ങൾക്ക് മുമ്പ് അവസാനത്തെ ഗ്ലേഷ്യൽ മാക്സിമത്തിന് ശേഷം ഫ്രാങ്കോ-കാൻ്റാബ്രിയൻ മേഖലയിലെ ഇൻട്രാ-യൂറോപ്യൻ വിപുലീകരണവുമായി H1, H3, കൂടാതെ സഹോദരി ഹാപ്ലോഗ് ഗ്രൂപ്പ് V എന്നിവയുടെ വ്യാപനം ബന്ധപ്പെട്ടിരിക്കുന്നുവെന്ന് അനുമാനിക്കപ്പെടുന്നു. പടിഞ്ഞാറൻ യൂറോപ്യൻ മൈറ്റോകോൺഡ്രിയൽ ഡിഎൻഎയുടെ ഗണ്യമായ അനുപാതം ഹാപ്ലോഗ്ഗ്രൂപ്പ് H1 ഉണ്ടാക്കുന്നു, കൂടാതെ ബാസ്കസ് ഓഫ് സ്പെയിൻ (28%), പോർച്ചുഗീസുകാർ (26%), അൻഡലൂഷ്യയിലെ നിവാസികൾ (24%), കാൻ്റബ്രിയയിലെ പാസിഗോ എത്നോഗ്രാഫിക് ഗ്രൂപ്പ് എന്നിവരും ഇത് പങ്കിടുന്നു. (24%). വടക്കേ ആഫ്രിക്കയിൽ, ലിബിയയിലെ ടുവാരെഗുകൾക്കിടയിൽ (61%) ഹാപ്ലോഗ് ഗ്രൂപ്പ് H1 ൻ്റെ അസാധാരണമായ ഉയർന്ന അനുപാതമുണ്ട്, ഇത് സ്പെയിനുമായി ചേർന്നുള്ള ഒരു പ്രദേശത്തെയും സൂചിപ്പിക്കുന്നു. കൂടാതെ, ഐബീരിയൻ പെനിൻസുല, വടക്കേ ആഫ്രിക്ക, സാർഡിനിയ എന്നിവിടങ്ങളിലെ മറ്റ് നിവാസികൾക്കിടയിൽ mtDNA ഹാപ്ലോഗ് ഗ്രൂപ്പ് H1 സാധാരണമാണ്. കൂടാതെ, ഫ്രാൻസ്, ബ്രിട്ടീഷ് ദ്വീപുകൾ, ആൽപ്സ്, കിഴക്കൻ യൂറോപ്പിലെ പല പ്രദേശങ്ങളിലും, യൂറോപ്പിലെ മറ്റ് സ്ഥലങ്ങളിൽ കുറഞ്ഞത് 5% വും അതിൻ്റെ വാഹകർ ജനസംഖ്യയുടെ 10% ആണ്. ഹാപ്ലോഗ് ഗ്രൂപ്പ് H1c യുടെ പ്രായം ഏകദേശം 9400 വർഷമാണ്. ഇക്കാര്യത്തിൽ, വടക്കൻ, കിഴക്കൻ യൂറോപ്പിലേക്കുള്ള അദ്ദേഹത്തിൻ്റെ നീക്കത്തിൻ്റെ സമയത്തെയും തീയതിയെയും കുറിച്ച് ചോദ്യം ഉയർന്നുവരുന്നു. 3500-2500 ബിസി സ്കാൻഡിനേവിയയിലെ ഫണൽ ബീക്കർ സംസ്കാരത്തിലെ കർഷകർക്കിടയിലാണ് ഈ ഉപവിഭാഗം കണ്ടെത്തിയത്. (ഗോഖേം2). സ്കാൻഡിനേവിയയിലെയും വടക്കൻ യൂറോപ്പിലെയും മുൻ സംസ്കാരങ്ങളിൽ സമാനമായ ഒരു ഹാപ്ലോടൈപ്പ് കണ്ടെത്തിയില്ല. യൂറോപ്പിൻ്റെ വടക്ക്, കിഴക്ക് ഭാഗങ്ങളിൽ ഇതിനകം മെസോ-നിയോലിത്തിക്ക്, ഒലെനി ദ്വീപിൽ നിന്ന് ഉക്രെയ്നിൻ്റെ തെക്ക് വരെയുള്ള വിശാലമായ സ്ട്രിപ്പിൽ ഈ ഉപവിഭാഗത്തിൻ്റെ രൂപം കണക്കിലെടുക്കുമ്പോൾ, തെക്ക് നിന്നുള്ള കുടിയേറ്റത്തിൻ്റെ ചോദ്യം ഉന്നയിക്കാൻ ഇത് ഞങ്ങളെ അനുവദിക്കുന്നു. മുൻ കാലഘട്ടത്തിൽ ഫ്രാങ്കോ-സ്പാനിഷ് പ്രദേശത്തെ ഈ ഹാപ്ലോഗ് ഗ്രൂപ്പിൻ്റെ വിതരണ കേന്ദ്രങ്ങളിലൊന്നായി കണക്കാക്കാമെന്നതിനാൽ, ഈ ഹാപ്ലോടൈപ്പിൻ്റെ രൂപത്തെക്കുറിച്ചുള്ള ഒരു യുക്തിസഹമായ അനുമാനം വടക്കൻ, കിഴക്കൻ യൂറോപ്പിലേക്ക് കടൽ വഴി കടന്നുകയറുന്നതിൻ്റെ പതിപ്പായി കണക്കാക്കാം. കാർഡിയാക് സെറാമിക്സ് സംസ്കാരത്തിൻ്റെ വാഹകരുടെ മെഡിറ്ററേനിയൻ തടം (നോർത്ത് പോണ്ടിക് ഇംപ്രെസോയെക്കുറിച്ചുള്ള ഡി. എൽ. ഗാസ്കെവിച്ചിൻ്റെ അനുമാനം). പടിഞ്ഞാറൻ യൂറോപ്പിന് പുറത്തുള്ള എച്ച് 1 കാരിയറുകളുടെ രൂപത്തെ കാർഡിയാക് സെറാമിക്സ് കോംപ്ലക്സിൻ്റെ മൈഗ്രേഷനുമായി (അല്ലെങ്കിൽ മറ്റ് സാംസ്കാരിക, പുരാവസ്തു ബന്ധങ്ങൾ) ബന്ധിപ്പിക്കാൻ ഇത് ഞങ്ങളെ അനുവദിക്കുന്നു, അതിൻ്റെ പ്രതിനിധികൾക്ക് ലെതർ ബോട്ടുകളിൽ തീരത്ത് സഞ്ചരിക്കാം. പടിഞ്ഞാറൻ യൂറോപ്പിൽ നിന്നും മെഡിറ്ററേനിയനിൽ നിന്നുമുള്ള മെഗാലിത്തിക് സംസ്കാരങ്ങളുടെയും കാർഡിയൽ സെറാമിക്സിൻ്റെയും സ്രഷ്ടാക്കളെ ഗാസ്കെവിച്ച് ഉദ്ധരിച്ചു, ആരുടെ പരിതസ്ഥിതിയിലാണ് ഈ ഉപവിഭാഗം കാണപ്പെടുന്നത്, ബിസി ആറാം സഹസ്രാബ്ദം വരെ നിലനിന്നിരുന്നു. ബഗ്-ഡൈനെസ്റ്റർ സംസ്കാരത്തിലൂടെ ക്രിമിയയിലേക്കും തെക്കൻ ഉക്രെയ്നിലേക്കും ബോസ്പോറസ് ഇസ്ത്മസ്.
വിശകലനത്തിൻ്റെ ഫലമായി, അവതരിപ്പിച്ച ഹാപ്ലോടൈപ്പ്, പല ഗവേഷകരും ഇന്തോ-യൂറോപ്യൻ എന്ന് കരുതുന്ന ഫണൽ ബീക്കർ സംസ്കാരത്തിൻ്റെ പ്രതിനിധിയുടെ ഗോഖേം 2 ൽ കാണപ്പെടുന്ന ഉപക്ലേഡുമായി പൊരുത്തപ്പെടുന്നുവെന്ന് നമുക്ക് പ്രസ്താവിക്കാം. മാതൃ ഹാപ്ലോഗ്രൂപ്പ് എച്ച്, പുരുഷ ഹാപ്ലോടൈപ്പുകളുടെ ആർ 1 എ 1 (സതേൺ ഒലെനി ഓസ്ട്രോവ്, സെർട്ടിയ) ൻ്റെ പുരാതന സാന്നിധ്യമുള്ള പ്രദേശത്താണ് കണ്ടെത്തിയത്, ഇത് റഷ്യൻ സമതലത്തിൽ നിന്നുള്ള ഈ ഹാപ്ലോഗ് ഗ്രൂപ്പിനെ പരിഗണിക്കാനും പുരുഷ R1a1 ൻ്റെ ഒരുതരം "കൂട്ടുകാരൻ" ആയി കണക്കാക്കാനും ഞങ്ങളെ അനുവദിക്കുന്നു. ദൈർഘ്യമേറിയ കണക്ഷനുകൾ H1c യുടെ വേരുകൾ മെഡിറ്ററേനിയനിലേക്ക് കണ്ടെത്തുന്നു. ഉപഭോക്താവിൻ്റെ വംശാവലിയെക്കുറിച്ചുള്ള വിശദമായ പഠനത്തിനായി, കൂടുതൽ വിശദമായ ടൈപ്പിംഗ് നടത്താൻ ശുപാർശ ചെയ്യുന്നു, കാരണം ഞങ്ങൾ നിങ്ങളെ ഓർമ്മിപ്പിക്കുന്നു, ഹാപ്ലോഗ് ഗ്രൂപ്പ് H1c-ൽ 20-ലധികം വ്യത്യസ്ത ഉപക്ലേഡുകൾ അടങ്ങിയിരിക്കുന്നു. അത്തരം ടൈപ്പിംഗ് ഉടൻ മോസ്കോ ലബോറട്ടറി ഓഫ് ഡിഎൻഎ വംശാവലി നടപ്പിലാക്കും. 15 ആയിരം വർഷങ്ങൾക്ക് മുമ്പ് തെക്കുകിഴക്കൻ യൂറോപ്പിലാണ് ഹാപ്ലോഗ് ഗ്രൂപ്പ് I2 ഉത്ഭവിച്ചത്. ബോസ്നിയ-ഹെർസഗോവിന, തെക്കൻ ഡാൽമേഷ്യ (ക്രൊയേഷ്യ) എന്നിവിടങ്ങളിലാണ് ഇതിൻ്റെ ഏറ്റവും ഉയർന്ന സാന്ദ്രത കാണപ്പെടുന്നത്, കൂടാതെ എല്ലാ തെക്കൻ സ്ലാവിക് ജനസംഖ്യയിലും ഗണ്യമായ ശതമാനമുണ്ട്.
ഈ മേഖലകളിലെ I2a യുടെ ഉയർന്ന സാന്ദ്രത സൂചിപ്പിക്കുന്നത്, ആധുനിക ക്രൊയേഷ്യയിലെ അഡ്രിയാറ്റിക് പ്രദേശവും ദിനാറിക് ആൽപ്സും കഴിഞ്ഞ ഹിമയുഗത്തിൽ I2a വഹിക്കുന്ന പാലിയോപോപ്പുലേഷനുകളുടെ സ്വാഭാവിക അഭയകേന്ദ്രമായിരുന്നു എന്നാണ്. ഹാപ്ലോഗ് ഗ്രൂപ്പ് I2a യുടെ പ്രതിനിധികൾ ബാൽക്കൻ പെനിൻസുലയിലെ ഏറ്റവും പുരാതന സംസ്കാരങ്ങളുടെ ഉത്ഭവസ്ഥാനത്ത് നിലകൊള്ളുന്നു: ചെൽ ക്ലഡോവ്, ലെപെൻസ്കി വീർ, ഗുര ബാസിയുലുയി, നികോമീഡിയ. ഈ വംശത്തിൻ്റെ അടിസ്ഥാനത്തിൽ, ഇല്ലിയറിയൻ, ത്രേസിയൻ ഗോത്രങ്ങൾ പ്രത്യക്ഷത്തിൽ വികസിച്ചു. I2a ബ്രാഞ്ച് സാർഡിനിയയിൽ ആദ്യമായി ജനപ്രീതി നേടിയ ഒന്നാണ്. ഹാപ്ലോഗ് ഗ്രൂപ്പ് ശാഖകൾ:I2a1ഹാപ്ലോഗ് ഗ്രൂപ്പ് I2a1(M26) സാർഡിനിയയിലെ എല്ലാ പുരുഷ വംശങ്ങളുടെയും ഏകദേശം 40% പ്രതിനിധീകരിക്കുന്നു. കൂടാതെ, ബാസ്ക് രാജ്യത്തും ഐബീരിയൻ പെനിൻസുലയിലും കുറഞ്ഞതും മിതമായതുമായ ആവൃത്തിയിലും ഇത് കാണപ്പെടുന്നു, കൂടാതെ മാസിഡോണിയയിൽ താമസിക്കുന്ന അൽബേനിയക്കാരുടെ ഒരു സാമ്പിളിൽ 1.6% (1/64) ഉം ഒരു സാമ്പിളിൽ 1.2% (3/257) ഉം ആണ്. ചെക്കോവ് M26 സബ്ക്ലേഡിനുള്ള YSTR വ്യതിയാനത്തിൻ്റെ പ്രായം 8.0 ± 4.0 ആയിരം വർഷമാണ് (റൂട്ട്സി 2004). ശാഖയാണെന്നാണ് അനുമാനം I2aനിന്നു മാറി നിന്നു I2ഏകദേശം 11,000 വർഷങ്ങൾക്ക് മുമ്പ് ബാൽക്കണിലും ഒരു ശാഖയും I2a1നിന്നു മാറി നിന്നു I2aഏകദേശം 8,000 വർഷങ്ങൾക്ക് മുമ്പ് സാർഡിനിയയിലോ അതിൻ്റെ സമീപത്തെ യൂറോപ്യൻ തീരത്ത് എവിടെയോ. I2a2 ഹാപ്ലോഗ് ഗ്രൂപ്പ് വിതരണം I2a2 I2a2(snp M423) തെക്ക്-കിഴക്കൻ യൂറോപ്പിലെ ജനസംഖ്യയുടെ സാധാരണമാണ്. ഡാൽമേഷ്യ (ക്രൊയേഷ്യ), ബോസ്നിയ-ഹെർസഗോവിന (>50%) എന്നിവിടങ്ങളിലാണ് ഏറ്റവും ഉയർന്ന സാന്ദ്രത. ദിനാറിക് ഹൈലാൻഡിലെ ഈ പ്രദേശത്താണ് ഈ ശാഖ ഉത്ഭവിച്ചതെന്ന് അനുമാനിക്കപ്പെടുന്നു I2aഏകദേശം 7500 വർഷങ്ങൾക്ക് മുമ്പ്. ഏറ്റവും സാധാരണമായ ഹാപ്ലോഗ് ഗ്രൂപ്പ് I2a2സ്ലാവിക് ജനതയിൽ (പ്രത്യേകിച്ച് തെക്കൻ സ്ലാവിക്) കാണപ്പെടുന്നു, കൂടാതെ റൊമാനിയക്കാർ, മോൾഡോവക്കാർ, ഹംഗേറിയക്കാർ, തെക്കൻ ലിത്വാനിയക്കാർ, അൽബേനിയക്കാർ, ഗ്രീക്കുകാർ, വടക്കുകിഴക്കൻ ഇറ്റലിയിലെ നിവാസികൾ, പടിഞ്ഞാറൻ അനറ്റോലിയ, വടക്കൻ കോക്കസസ് എന്നിവിടങ്ങളിൽ കാണപ്പെടുന്നു. ഹാപ്ലോഗ് ഗ്രൂപ്പിൻ്റെ ഏറ്റവും ഉയർന്ന ആവൃത്തിയും വൈവിധ്യവും I2a2പടിഞ്ഞാറൻ ബാൽക്കണിലെ ജനസംഖ്യയിൽ ആധുനിക ക്രൊയേഷ്യയിലെ അഡ്രിയാറ്റിക് പ്രദേശം അവസാന ഹിമയുഗത്തിൽ ഈ ഹാപ്ലോഗ് ഗ്രൂപ്പിൻ്റെ വാഹകർക്ക് അഭയകേന്ദ്രമായി പ്രവർത്തിച്ചതായി സൂചിപ്പിക്കുന്നു. Snp P37.2 ൻ്റെ ശാഖ ഏകദേശം 10.7 ± 4.8 ആയിരം വർഷങ്ങൾക്ക് മുമ്പ് സംഭവിച്ചു (Rootsi 2004). snp P37.2 ശാഖയുടെ പ്രായം 8.0 ± 4.0 ആയിരം വർഷമാണ് (Rootsi 2004), snp M423 ന് ഇത് 8.8 ± 3.6 ആയിരം വർഷമാണ് (അണ്ടർഹിൽ 2007). ഈ ശാഖയുടെ വ്യാപനം "ഹോളോസീൻ പരിവർത്തനത്തിന് മുമ്പല്ല, ആദ്യകാല നവീന ശിലായുഗത്തിന് ശേഷമല്ല" (പെരിസിക് 2005) സംഭവിച്ചതെന്ന് പെരിസിക് വിശ്വസിക്കുന്നു. ഏകാഗ്രതയുടെ രണ്ടാമത്തെ കേന്ദ്രം I2a2ഇന്നത്തെ മോൾഡോവയിൽ സ്ഥിതി ചെയ്യുന്നതും ട്രിപ്പിലിയൻ സംസ്കാരത്തിൻ്റെ അതിരുകളുമായി ഏകദേശം ഒത്തുപോകുന്നതും, പ്രത്യക്ഷത്തിൽ, ഈ സബ്ഹാപ്ലോഗ്രൂപ്പ് ഈ സംസ്കാരത്തിൻ്റെ സവിശേഷതയായിരുന്നു, കൂടാതെ ഇന്തോ-യൂറോപ്യൻമാരുടെ വരവോടെ, മറ്റ് ശാഖകളെപ്പോലെ അത്തരം അടിച്ചമർത്തൽ അനുഭവിക്കാതെ അവരുടെ ജീൻ പൂളിൽ ചേർന്നു. ഹാപ്ലോഗ് ഗ്രൂപ്പ് ഐമധ്യ, പടിഞ്ഞാറൻ യൂറോപ്പിൽ. I2b1സബ്പ്ലോഗ്രൂപ്പ് വിതരണങ്ങൾ I2b1ഹാപ്ലോഗ് ഗ്രൂപ്പിൻ്റെ വിതരണവുമായി പൊരുത്തപ്പെടുന്നു I1, ഫെനോസ്കാൻഡിയ ഒഴികെ, ഹാപ്ലോഗ് ഗ്രൂപ്പും സ്ഥിതിചെയ്യുന്ന പാലിയോലിത്തിക്ക് ഷെൽട്ടറുകളിലൊന്നിലെങ്കിലും അതിൻ്റെ സാന്നിധ്യം നിർദ്ദേശിക്കുന്നു. I1. അഭാവം I2b1 Fennoscandia ൽ ഹാപ്ലോഗ്ഗ്രൂപ്പ് എന്ന വസ്തുത കാരണം ആയിരിക്കാം I2b1ഈ പ്രദേശത്തെ ആദ്യകാല വാസസ്ഥലങ്ങൾ "സ്ഥാപക പ്രഭാവവും" ജനിതക വ്യതിയാനവും സ്വാധീനിക്കപ്പെട്ടു, കാരണം ഹാപ്ലോഗ് ഗ്രൂപ്പ് മുതൽ I2b1ലോവർ സാക്സോണിക്ക് പുറത്തുള്ള നിർദ്ദിഷ്ട പ്രദേശത്തെ ജനസംഖ്യയുടെ മൊത്തം Y-ക്രോമസോം വൈവിധ്യത്തിൻ്റെ 10% ൽ താഴെയാണ്. ഹാപ്ലോഗ് ഗ്രൂപ്പുകളുടെ വിതരണം I1ഒപ്പം I2b1ജർമ്മനിക് ഭാഷകൾ സംസാരിക്കുന്നവരുടെ വിതരണത്തിൻ്റെ ചരിത്രപരമായ അതിരുകളുമായി താരതമ്യേന നന്നായി ബന്ധപ്പെട്ടിരിക്കുന്നു. ഹാപ്ലോഗ് ഗ്രൂപ്പ് I2b1ജർമ്മനി, നെതർലാൻഡ്സ്, ബെൽജിയം, ഡെൻമാർക്ക്, ഇംഗ്ലണ്ട് (വെയിൽസ്, കോൺവാൾ ഒഴികെ), സ്കോട്ട്ലൻഡ്, സ്വീഡൻ്റെയും നോർവേയുടെയും തെക്കേ അറ്റം, അതുപോലെ നോർമാണ്ടി, മെയ്ൻ, അൻജൗ എന്നീ പ്രവിശ്യകളിൽ മാത്രം ജനസംഖ്യയുടെ 4% ൽ കൂടുതലായി കാണപ്പെടുന്നു. വടക്കുപടിഞ്ഞാറൻ ഫ്രാൻസിലെ പെർഷെ, തെക്കുകിഴക്കൻ ഫ്രാൻസിലെ പ്രോവൻസിൽ, ഇറ്റലിയിലെ ചരിത്ര പ്രദേശങ്ങളിൽ - ടസ്കാനി, ഉംബ്രിയ, ലാറ്റിയം; അതുപോലെ മോൾഡോവയിലും റിയാസാൻ മേഖലയിലും മൊർഡോവിയയിലും. ആധുനിക ഫ്രാൻസ്, ഇംഗ്ലണ്ട്, ഇറ്റലി, കിഴക്കൻ യൂറോപ്പ് എന്നിവിടങ്ങളിൽ I1, I2b1 എന്നിവയുടെ സാന്നിധ്യം ഇതിനകം തന്നെ കെൽറ്റിക്, ജർമ്മനിക് വികാസവുമായി ബന്ധപ്പെട്ടിരിക്കുന്നുവെന്ന് തോന്നുന്നു, ഇൻഡോ-യൂറോപ്യൻ കാലഘട്ടത്തിന് മുമ്പുള്ള ഈ ഹാപ്ലോഗ് ഗ്രൂപ്പുകൾ വടക്കൻ പ്രദേശങ്ങളിൽ മാത്രം കേന്ദ്രീകരിച്ചിരുന്നു. യൂറോപ്പ്. ഹാപ്ലോഗ് ഗ്രൂപ്പിൻ്റെ ശാഖകളിലൊന്ന് I2b1, അതായത് I2b1a(snp M284), ബ്രിട്ടീഷ് ജനസംഖ്യയിൽ മാത്രം കാണപ്പെടുന്നു, ഇത് ബ്രിട്ടീഷ് ദ്വീപുകളിൽ അതിൻ്റെ നീണ്ട ചരിത്രത്തെ സൂചിപ്പിക്കാം. രസകരമെന്നു പറയട്ടെ, ഹാപ്ലോഗ് ഗ്രൂപ്പിൻ്റെ കുറഞ്ഞ ആവൃത്തിയിൽ I1ഒപ്പം I2bആധുനിക തുർക്കിയിലെ ബിഥീനിയ, ഗലാത്തിയ എന്നീ ചരിത്ര പ്രദേശങ്ങളിൽ ഇവ കാണപ്പെടുന്നു, അവിടെ ബിഥീനിയയിലെ നിക്കോമിഡെസ് ഒന്നാമൻ്റെ ക്ഷണപ്രകാരം കുടിയേറിയ കെൽറ്റുകൾക്ക് അവ കൊണ്ടുവരാമായിരുന്നു. ഹാപ്ലോഗ് ഗ്രൂപ്പ് I2b1സാർഡിനിയൻ ജനസംഖ്യയുടെ ഏകദേശം 1% ലും ഇത് സംഭവിക്കുന്നു. എന്നാണ് അനുമാനിക്കുന്നത് I2bനിന്ന് വേറിട്ട് നിൽക്കുന്നു I2മധ്യ യൂറോപ്പിൽ, ഏകദേശം 13 ആയിരം വർഷങ്ങൾക്ക് മുമ്പ്, പതുക്കെ പിൻവാങ്ങുന്ന ഹിമാനിയുടെ അരികിൽ, I2b1- നിന്ന് I2bകൂടുതൽ വടക്ക്, ഇപ്പോൾ ജർമ്മനിയിൽ, ഏകദേശം 9 ആയിരം വർഷങ്ങൾക്ക് മുമ്പ്. പ്രത്യേകിച്ച് ബ്രിട്ടീഷ് ബ്രാഞ്ച് I2b1aനിന്നു മാറി നിന്നു I2b1ഏകദേശം 3 സഹസ്രാബ്ദങ്ങൾക്ക് മുമ്പ്. I2b2മധ്യ ജർമ്മനിയിലെ വെങ്കലയുഗത്തിലെ പുരാവസ്തു സൈറ്റായ ലിച്ചെൻസ്റ്റൈൻ ഗുഹയിൽ കണ്ടെത്തിയ അസ്ഥികൂട അവശിഷ്ടങ്ങളിൽ നിന്നാണ് ഹാപ്ലോഗ് ഗ്രൂപ്പ് I2b2 കണ്ടെത്തിയത്, അവിടെ ഉർൺ ഫീൽഡ് സംസ്കാരത്തിൽ നിന്നുള്ള പുരാവസ്തുക്കളും കണ്ടെത്തി. ഗുഹയിലെ 19 പുരുഷന്മാരുടെ അവശിഷ്ടങ്ങളിൽ, 13 ൽ ഹാപ്ലോഗ് ഗ്രൂപ്പ് I2b2, ഒന്നിൽ R1b, രണ്ടിൽ R1a എന്നിവ കണ്ടെത്തി. ഹാപ്ലോഗ് ഗ്രൂപ്പ് I2b യുടെ അന്നത്തെ വ്യാപനത്തിൻ്റെ പ്രഭവകേന്ദ്രത്തിലാണ് ഗുഹ സ്ഥിതിചെയ്യുന്നത്. മുൻ യുഗോസ്ലാവിയ, റൊമാനിയ, ബൾഗേറിയ, സാർഡിനിയ എന്നിവിടങ്ങളിലെ ഏറ്റവും സാധാരണമായ പിതൃപരമ്പരയാണ് ഹാപ്ലോഗ് ഗ്രൂപ്പ് I2, കൂടാതെ മിക്ക സ്ലാവിക് രാജ്യങ്ങളിലെയും ഒരു പ്രധാന വംശപരമ്പരയാണ്. ബോസ്നിയയിൽ (55%, ബോസ്നിയൻ ക്രൊയറ്റുകളിൽ 71% ഉൾപ്പെടെ), സാർഡിനിയ (39.5%), ക്രൊയേഷ്യ (38%), സെർബിയ (33%), മോണ്ടിനെഗ്രോ (31%), റൊമാനിയ (28%), മോൾഡോവ എന്നിവിടങ്ങളിൽ ഇതിൻ്റെ പരമാവധി ആവൃത്തികൾ നിരീക്ഷിക്കപ്പെടുന്നു. (24%), മാസിഡോണിയ (24%), സ്ലൊവേനിയ (22%), ബൾഗേറിയ (22%), ബെലാറസ് (18.5%), ഹംഗറി (18%), സ്ലൊവാക്യ (17.5%), ഉക്രെയ്ൻ (13.5%), അൽബേനിയ ( 13.5%). ജർമ്മൻ രാജ്യങ്ങളിൽ ഇത് 5 മുതൽ 10% വരെ ആവൃത്തിയിൽ കാണപ്പെടുന്നു. I2 ൻ്റെ ഫൈലോജെനിനിങ്ങൾ ജനിതക വംശാവലിയിൽ പുതിയ ആളാണെങ്കിൽ, ഒരു ഫൈലോജെനെറ്റിക് ട്രീ എങ്ങനെ വായിക്കാമെന്ന് മനസിലാക്കാൻ ഫൈലോജെനെറ്റിക്സിലേക്കുള്ള ഞങ്ങളുടെ ആമുഖം പരിശോധിക്കുക. I2 ൻ്റെ ഫൈലോജെനെറ്റിക് ട്രീ കഴിഞ്ഞ 10 വർഷമായി വളരെയധികം വികസിച്ചു, കൂടാതെ R1b ഒഴികെയുള്ള മറ്റേതൊരു ഹാപ്ലോഗ് ഗ്രൂപ്പിനേക്കാളും ഉയർന്ന തലത്തിലുള്ള സബ്ക്ലേഡുകൾ പുനർനാമകരണം ചെയ്യപ്പെട്ടു. എല്ലാ ആശയക്കുഴപ്പങ്ങളും ഒഴിവാക്കാൻ, പഴയ പഠനങ്ങൾ വായിക്കുമ്പോഴോ ഒരു പ്രത്യേക സബ്ക്ലേഡിലേക്ക് പരാമർശം നടത്തുമ്പോഴോ നിർവചിക്കുന്ന മ്യൂട്ടേഷനുകൾ (എസ്എൻപികൾ) രണ്ടുതവണ പരിശോധിക്കാൻ ശുപാർശ ചെയ്യുന്നു.സ്പേസ് കുറവായതിനാൽ നിരവധി ചെറിയ സബ്ക്ലേഡുകൾ ഉൾപ്പെടുത്തിയിട്ടില്ല.  വലുതാക്കാൻ ക്ലിക്ക് ചെയ്യുക
ഉത്ഭവവും ചരിത്രവുംഹാപ്ലോഗ് ഗ്രൂപ്പ് I യൂറോപ്പിലെ ഏറ്റവും പഴക്കമേറിയ ഹാപ്ലോഗ് ഗ്രൂപ്പാണ്, എല്ലാ സാധ്യതയിലും അവിടെ ഉത്ഭവിച്ച ഒരേയൊരു ഗ്രൂപ്പാണ് (C1a2 പോലുള്ള വളരെ ചെറിയ ഹാപ്ലോഗ് ഗ്രൂപ്പുകളും മറ്റ് ഹാപ്ലോഗ് ഗ്രൂപ്പുകളുടെ ആഴത്തിലുള്ള ഉപവിഭാഗങ്ങളും ഒഴികെ). ഏകദേശം 35,000 വർഷങ്ങൾക്ക് മുമ്പ് ഹാപ്ലോഗ് ഗ്രൂപ്പ് IJ മിഡിൽ ഈസ്റ്റിൽ നിന്ന് യൂറോപ്പിലേക്ക് എത്തുമായിരുന്നു, പിന്നീട് ഹാപ്ലോഗ് ഗ്രൂപ്പ് I ആയി വികസിച്ചു. ഔറിഗ്നേഷ്യൻ കാലഘട്ടത്തിൽ (45,000 മുതൽ 28,000 വർഷങ്ങൾക്ക് മുമ്പ്) യൂറോപ്പിനെ കോളനിവത്കരിച്ച ആദ്യത്തെ ഹോമോ സാപ്പിയൻസ് CT, C1a, C1b, F, I എന്നീ ഹാപ്ലോഗ് ഗ്രൂപ്പുകളിൽ പെട്ടവരാണെന്ന് ഇപ്പോൾ സ്ഥിരീകരിച്ചിട്ടുണ്ട്. I2 (M438/P215/S31) 26,500 മുതൽ 19,000 വർഷങ്ങൾക്ക് മുമ്പ് നിലനിന്നിരുന്ന ലാസ്റ്റ് ഗ്ലേഷ്യൽ മാക്സിമം (LGM) കാലഘട്ടത്തിൽ, അവസാനത്തെ പാലിയോലിത്തിക്ക് കാലഘട്ടത്തിൽ ഉത്ഭവിച്ചതായി കരുതപ്പെടുന്നു. പാലിയോലിത്തിക്ക് യൂറോപ്യന്മാർ നാടോടികളായ വേട്ടയാടുന്നവരായതിനാൽ അതിൻ്റെ കൃത്യമായ ഉത്ഭവ പ്രദേശം നിർണ്ണയിക്കാൻ കഴിയില്ലെങ്കിലും പടിഞ്ഞാറൻ യൂറോപ്പിലാണ് I2 പ്രത്യക്ഷപ്പെട്ടത്. പുരാവസ്തു അസ്ഥികൂടങ്ങളിൽ നിന്ന് കണ്ടെടുത്ത ഏറ്റവും പഴയ I2 സാമ്പിൾ, അസീലിയൻ സംസ്കാരവുമായി ബന്ധപ്പെട്ട സ്വിറ്റ്സർലൻഡിലെ ഗ്രോട്ടെ ഡു ബിച്ചോണിൽ നിന്നുള്ള 13,500 വർഷം പഴക്കമുള്ള ആളാണ് (ജോൺസ് et al. (2015) കാണുക). അവൻ്റെ മാതൃപരമ്പര U5b1h ആയിരുന്നു. 2016 ഒക്ടോബർ വരെ, 15 മെസോലിത്തിക്ക് യൂറോപ്യൻ Y-DNA സാമ്പിളുകൾ പരീക്ഷിച്ചു. അവരിൽ ഒരാൾ ഹാപ്ലോഗ് ഗ്രൂപ്പ് C1a2 (സ്പെയിനിൽ), ഒന്ന് F (ജർമ്മനിയിൽ), രണ്ട് I* (ഫ്രാൻസിൽ), ആറ് I2 (ലക്സംബർഗ്, സ്വീഡൻ) എന്നിവയിൽ ഉൾപ്പെടുന്നു, I2a1* (P37.2), I2a1a1a ഉൾപ്പെടെ. (L672), I2a1b (M423), I2c2 (PF3827). U2e, U4, U5a1, U5a2, U5b എന്നിവയായിരുന്നു അവർ വഹിച്ച മാതൃ (mtDNA) വംശങ്ങൾ. റഷ്യയിൽ നിന്നുള്ള നാല് സാമ്പിളുകൾ Y-haplogroup J*, R1a1 (2x), R1b1a എന്നിവയുടേതാണ്. മെസോലിത്തിക്ക് യൂറോപ്യൻ വംശങ്ങൾക്കിടയിൽ ഇതിനകം ഒരു നിശ്ചിത അളവിലുള്ള വൈവിധ്യം ഉണ്ടായിരുന്നുവെന്ന് ഇത് കാണിക്കുന്നു, എന്നിരുന്നാലും ഈ വംശങ്ങളിൽ പലതും (C1a2, F, I*, J*) ഇപ്പോൾ വളരെ വിരളമാണ്. ഹാപ്ലോഗ് ഗ്രൂപ്പ് I2a1 നിയോലിത്തിക്ക് കാലഘട്ടത്തിൽ നിന്ന് വ്യക്തമല്ലാത്ത കാരണങ്ങളാൽ വലിയ വിജയിയായി മാറിയതായി തോന്നുന്നു. മാത്തിസൺ മറ്റുള്ളവരുടെ അനറ്റോലിയയിൽ നിന്നുള്ള ആദ്യകാല നിയോലിത്തിക്ക് ജീനോമുകളുടെ വിശകലനം. (2015) കാണിക്കുന്നത് I2c (L596) 8,500 വർഷങ്ങൾക്ക് മുമ്പ് വടക്ക്-പടിഞ്ഞാറൻ തുർക്കിയിൽ ഉണ്ടായിരുന്നുവെന്നും ഫലഭൂയിഷ്ഠമായ ചന്ദ്രക്കലയിൽ നിന്ന് യൂറോപ്പിലേക്കുള്ള യാത്രാമധ്യേ നിയോലിത്തിക്ക് കർഷകർ ഇത് സ്വാംശീകരിച്ചിരുന്നുവെന്നും. 2014-നും 2016-നും ഇടയിൽ നടത്തിയ വിവിധ പഠനങ്ങൾ, മധ്യശിലായുഗത്തിലെ എല്ലാ യൂറോപ്യൻ വംശങ്ങളും നിയോലിത്തിക്ക് കർഷകരുടെ തരംഗത്താൽ ആഗിരണം ചെയ്യപ്പെട്ടതായി വെളിപ്പെടുത്തി. എങ്കിലും I2a അവയിൽ ഏറ്റവും സാധാരണമായത്, പരിശോധിച്ച 69 നിയോലിത്തിക്ക് Y-DNA സാമ്പിളുകളിൽ 16% ആണ്. അനറ്റോലിയൻ കർഷകരുടെ യഥാർത്ഥ വംശപരമ്പരയായ G2a കഴിഞ്ഞാൽ ഏറ്റവും സാധാരണമായ രണ്ടാമത്തെ പിതൃപരമ്പരയായി ഇത് സ്ഥാപിക്കുന്നു. I2a1 തെക്കുകിഴക്കൻ യൂറോപ്പിലെ Starčevo-Kőrсs-Criş സംസ്കാരത്തിൽ (ബിസി 6000-4500) പ്രത്യേകിച്ച് അഭിവൃദ്ധി പ്രാപിച്ചതായി തോന്നുന്നു, അവിടെ അത് ഇന്നും പ്രധാന പുരുഷ വംശപരമ്പരകളിൽ ഒന്നാണ്. പടിഞ്ഞാറൻ മെഡിറ്ററേനിയനിലെ അച്ചടിച്ച-കാർഡിയം മൺപാത്ര സംസ്കാരത്തിലും (ബിസി 5000-1500) I2a1 പ്രത്യക്ഷപ്പെട്ടു. ആധുനിക സാർഡിനിയക്കാരും ബാസ്കുകളും അവരുടെ ജീനോമിൻ്റെ ഏറ്റവും വലിയ പങ്ക് ആ നിയോലിത്തിക്ക് കർഷകരിൽ നിന്ന് അച്ചടിച്ച-കാർഡിയം പോട്ടറി സംസ്കാരത്തിൽ നിന്ന് പാരമ്പര്യമായി സ്വീകരിച്ചു. ആദ്യകാല അല്ലെങ്കിൽ മധ്യ നിയോലിത്തിക്ക് സാമ്പിളുകളിൽ ഇതുവരെ ഒരു I2a2 മാത്രമേ കണ്ടെത്തിയിട്ടുള്ളൂവെങ്കിലും (സ്പെയിനിൽ നിന്നുള്ള ഒരു I2a2a-M223), അവയിൽ പലതും ചെമ്പ്, വെങ്കല യുഗങ്ങളിൽ കാണപ്പെട്ടു. ചെമ്പ്, വെങ്കലയുഗ സാമ്പിളുകളിൽ സ്പെയിനിൽ ഒരു I2a2, രണ്ട് I2a2a, ഒരു I2a2a1 (CTS616), തെക്കൻ റഷ്യയിൽ ഒരു I2a2a1b1b2 (S12195) (Yamna culture), ഒരു I2a2a1 (CTS9183), ഒരു I2a2a1 (CTS9183) (H2a2a2a2a) സംസ്കാരം എന്നിവ ഉൾപ്പെടുന്നു. ), ജർമ്മനിയിൽ ആറ് I2a2b (L38) (Unetice and Urnfield cultures). ചെമ്പ് യുഗത്തിൽ നിന്നുള്ള I2a1 സാമ്പിളുകളിൽ വടക്കൻ ഇറ്റലിയിൽ നിന്നുള്ള I2a1a1 (റെമെഡെല്ലോ സംസ്കാരം), ഹംഗറിയിൽ നിന്നുള്ള I2a1 (വാത്യ സംസ്കാരം) ഉൾപ്പെടുന്നു. ജർമ്മനിയിലെ Unetic സംസ്കാരത്തിലും I2c2 കണ്ടെത്തി. ഹാപ്ലോഗ് ഗ്രൂപ്പ് I2a1 (P37.2)ഹാപ്ലോഗ് ഗ്രൂപ്പ് I2a1 ആണ് I2 ൻ്റെ ഏറ്റവും വലിയ ശാഖയും തെക്ക്-കിഴക്ക്, തെക്ക്-പടിഞ്ഞാറ്, വടക്ക്-പടിഞ്ഞാറ് യൂറോപ്പിലെ നിയോലിത്തിക്ക് സംസ്കാരങ്ങളുമായി ഏറ്റവും ശക്തമായി ബന്ധപ്പെട്ടിരിക്കുന്നതും.  ഹാപ്ലോഗ് ഗ്രൂപ്പ് I2a1a1 (M26)I2a1a (M26, L158, L159.1/S169.1) 2005 വരെ I1b2 എന്നും 2006-7 ൽ I1b1b എന്നും 2008 മുതൽ 2010 വരെ I2a1 എന്നും അറിയപ്പെട്ടിരുന്നു. ഇത് എല്ലാ പടിഞ്ഞാറൻ യൂറോപ്പിലും കാണപ്പെടുന്നു, കൂടാതെ സാർഡിനിയക്കാർക്കിടയിൽ പരമാവധി ആവൃത്തിയിൽ എത്തുന്നു (37.5%) ബാസ്ക്വസ് (5%), രണ്ട് ജനസംഖ്യാ ഒറ്റപ്പെടലുകൾ. M26 ഭൂമിശാസ്ത്രപരമായി ബ്രിട്ടീഷ് ദ്വീപുകൾ, താഴ്ന്ന രാജ്യങ്ങൾ, ഫ്രാൻസ്, പടിഞ്ഞാറൻ ജർമ്മനി, സ്വിറ്റ്സർലൻഡ്, സാർഡിനിയ, സിസിലി, ഇറ്റലിയുടെ പടിഞ്ഞാറൻ തീരം, ഐബീരിയ, മഗ്രിബിൻ്റെ മെഡിറ്ററേനിയൻ തീരം എന്നിവിടങ്ങളിൽ പരിമിതപ്പെടുത്തിയിരിക്കുന്നു. L160 മ്യൂട്ടേഷനുള്ള ഒരേയൊരു M26 നെഗറ്റീവ് അയർലണ്ടിൽ മാത്രം ഒതുങ്ങുന്നു. I2a1a-M26 നിയോലിത്തിക്ക്, ചാൽക്കോലിത്തിക്ക് കാലഘട്ടങ്ങളിൽ പടിഞ്ഞാറൻ യൂറോപ്പിലെ മെഗാലിത്തിക് സംസ്കാരങ്ങളിലെ പ്രധാന പിതൃപരമ്പരകളിൽ ഒന്നായിരിക്കാം. I2a1a1a (L672) ഇതിനകം തന്നെ മെസോലിത്തിക് സ്വീഡനിൽ കണ്ടെത്തിയിരുന്നു, ഇത് സൂചിപ്പിക്കുന്നത് I2a1a മധ്യശിലായുഗ കാലഘട്ടത്തിൽ ഐബീരിയ മുതൽ സ്കാൻഡിനേവിയ വരെ വളരെ വ്യാപകമായ വ്യാപനം ഉണ്ടായിരുന്നു എന്നാണ്. പിന്നീട്, സമീപ കിഴക്കൻ പുതുമുഖങ്ങളുമായി ഇടകലർന്ന് അവർ കൃഷി സ്വീകരിക്കുമായിരുന്നു. ഹാപ്ലോഗ് ഗ്രൂപ്പ് I2a1b (M423)I2a1b (M423, L178) 2007 വരെ I1b എന്നും 2008 മുതൽ 2010 വരെ I2a2 എന്നും അറിയപ്പെട്ടിരുന്നു. പ്രധാന ഉപവിഭാഗം, എല്ലാ M423 വംശാവലികളിൽ 90%-ലധികവും പ്രതിനിധീകരിക്കുന്നത് L621അതിൻ്റെ ഉപവിഭാഗവും L147.2. എന്നിവയാണ് മറ്റ് ഉപവിഭാഗങ്ങൾ L41.2(വളരെ അപൂർവ്വം) കൂടാതെ L161.1(കൂടുതലും ജർമ്മനിയിലും ബ്രിട്ടീഷ് ദ്വീപുകളിലും കാണപ്പെടുന്നു). ഹാപ്ലോഗ് ഗ്രൂപ്പ് I2a1b-L621ഈ ശാഖ സ്ലാവിക് രാജ്യങ്ങളിൽ കൂടുതലായി കാണപ്പെടുന്നു. ദിനാറിക് സ്ലാവുകൾക്കിടയിലും (സ്ലോവേനികൾ, ക്രൊയേഷ്യക്കാർ, ബോസ്നിയാക്കുകൾ, സെർബുകൾ, മോണ്ടിനെഗ്രിൻസ്, മാസിഡോണിയക്കാർ) അതുപോലെ ബൾഗേറിയ, റൊമാനിയ, മോൾഡേവിയ, പടിഞ്ഞാറൻ ഉക്രെയ്ൻ, ബെലാറസ് എന്നിവിടങ്ങളിൽ ഇതിൻ്റെ പരമാവധി ആവൃത്തികൾ നിരീക്ഷിക്കപ്പെടുന്നു. അൽബേനിയ, ഗ്രീസ്, ഹംഗറി, സ്ലൊവാക്യ, പോളണ്ട്, തെക്ക്-പടിഞ്ഞാറൻ റഷ്യ എന്നിവിടങ്ങളിലും ഇത് സാധാരണമാണ്. I2-L621 (L147.2+) എന്നും അറിയപ്പെടുന്നു I2a-Din(ദിനറിക്കിന്). വടക്കുകിഴക്കൻ റൊമാനിയ, മോൾഡോവ, മധ്യ ഉക്രെയ്ൻ എന്നിവിടങ്ങളിൽ I2a1b-L621 ൻ്റെ ഉയർന്ന സാന്ദ്രത കുക്കുട്ടേനി-ട്രിപ്പിലിയൻ സംസ്കാരത്തിൻ്റെ (ബിസി 4800-3000) പരമാവധി വ്യാപനത്തെ ഓർമ്മിപ്പിക്കുന്നു. ഈ സംസ്കാരത്തിൽ നിന്നുള്ള Y-DNA സാമ്പിളുകളൊന്നും ഇന്നുവരെ പരീക്ഷിച്ചിട്ടില്ല, എന്നാൽ ഇത് Starčevo-Kőrсs-Criş സംസ്കാരത്തിൽ നിന്നുള്ള ഒരു ശാഖയായി പരിണമിച്ചതിനാൽ, I2a അതിൻ്റെ പ്രധാന പിതൃപരമ്പരകളിൽ ഒന്നായിരിക്കാം, കൂടാതെ ഒരു സ്ഥാപക പ്രഭാവം വർദ്ധിക്കുകയും ചെയ്തേക്കാം. അതിൻ്റെ ആവൃത്തി വെങ്കലയുഗത്തിലെ ഇന്തോ-യൂറോപ്യൻ അധിനിവേശത്തിന് മുമ്പ് യൂറോപ്പിലെ ഏറ്റവും പുരോഗമിച്ച നവീന ശിലായുഗ സംസ്കാരമായിരുന്നു കുക്കുട്ടേനി-ട്രിപ്പിലിയൻ സംസ്കാരം, യമ്ന ബാൽക്കണിലേക്കും മധ്യ യൂറോപ്പിലേക്കും വ്യാപിക്കുന്നതിന് മുമ്പ് സ്റ്റെപ്പി സംസ്കാരവുമായി തീവ്രമായ ബന്ധങ്ങൾ ഉണ്ടായിരുന്നതായി തോന്നുന്നു (R1a യുടെ ചരിത്രങ്ങൾ കാണുക. കൂടാതെ R1b). ബിസി 3500 മുതൽ, പോണ്ടിക്-കാസ്പിയൻ സ്റ്റെപ്പിലെ യമ്ന കാലഘട്ടത്തിൻ്റെ ആരംഭത്തിൽ, കുക്കുട്ടേനി-ട്രിപ്പിലിയൻ ജനത കിഴക്ക് ഇന്നത്തെ പടിഞ്ഞാറൻ ഉക്രെയ്നിൻ്റെ സ്റ്റെപ്പിയിലേക്ക് വ്യാപിക്കാൻ തുടങ്ങി, അവരുടെ പട്ടണങ്ങൾ (അക്കാലത്ത് ലോകത്തിലെ ഏറ്റവും വലിയത്) അവരുടെ യമ്ന അയൽക്കാരെപ്പോലെ വർദ്ധിച്ചുവരുന്ന നാടോടികളായ ജീവിതശൈലി സ്വീകരിക്കുകയും ചെയ്യുന്നു. കുക്കുട്ടേനി-ട്രിപ്പിലിയൻ ജനതയെ യമ്ന അയൽക്കാർ സ്വാംശീകരിച്ചുവെന്നും അവർ കോർഡഡ് വെയർ വിപുലീകരണത്തോടെ ബാൾട്ടിക്കിലേക്ക് മുന്നേറുമ്പോൾ ഹാപ്ലോഗ് ഗ്രൂപ്പുകളായ R1a, R1b എന്നിവയ്ക്കൊപ്പം ഒരു ന്യൂനപക്ഷ വംശമായി വ്യാപിച്ചുവെന്നും എളുപ്പത്തിൽ സങ്കൽപ്പിക്കാൻ കഴിയും. പകരമായി, I2-L621 വംശജർക്ക് മുഖ്യധാരാ പ്രോട്ടോ-ഇന്തോ-യൂറോപ്യൻ സമൂഹത്തിൽ നിന്ന് ആപേക്ഷികമായി ഒറ്റപ്പെട്ട് ജീവിക്കാമായിരുന്നു, ഉക്രെയ്നിലോ പോളണ്ടിലോ ബെലാറസിലോ എവിടെയെങ്കിലും, നൂറ്റാണ്ടുകളും സഹസ്രാബ്ദങ്ങളും കടന്നുപോകുമ്പോൾ, അവർക്ക് ചുറ്റുമുള്ള പ്രധാനമായും R1a ജനസംഖ്യയുമായി കൂടിച്ചേരുമായിരുന്നു. തത്ഫലമായുണ്ടാകുന്ന സംയോജനം പ്രോട്ടോ-സ്ലാവുകളുടെ പൂർവ്വികർ ആകുമായിരുന്നു. ഇക്കാലത്ത്, തെക്കുകിഴക്കൻ യൂറോപ്പിൽ G2a-യേക്കാൾ അഞ്ച് മുതൽ പത്തിരട്ടി വരെ കൂടുതലാണ് I2a1, അതേസമയം നിയോലിത്തിക്ക് കാലഘട്ടത്തിൽ G2a ഏകദേശം നാലിരട്ടി കൂടുതലായിരുന്നു. ഈ പൂർണ്ണമായ വിപരീതത്തെ എന്താണ് വിശദീകരിക്കാൻ കഴിയുക? ചരിത്രത്തിലെ ഒരു ഘട്ടത്തിൽ, I2a1 വംശജർ വിജയിക്കുന്ന ഭാഗത്ത് നിന്ന് പ്രയോജനം നേടിയതായി തോന്നുന്നു. യൂറോപ്പിലേക്കുള്ള യമ്നയുടെ പടിഞ്ഞാറൻ വിപുലീകരണത്തിൽ (സാങ്കൽപ്പികമായി) ചേരുന്നതിൽ നിന്നുള്ള ഒരു ചെറിയ ഉത്തേജനം കൂടാതെ, I2a1b-L621 ഒരു പ്രധാന കിഴക്കൻ യൂറോപ്യൻ വംശാവലിയായി മാറാൻ അനുവദിച്ച പ്രധാന നിർണ്ണായക സംഭവം 6-ആം നൂറ്റാണ്ട് മുതൽ 9-ആം നൂറ്റാണ്ട് വരെയുള്ള സ്ലാവിക് കുടിയേറ്റമാണ്. മിക്ക ആധുനിക കിഴക്കൻ യൂറോപ്യന്മാരും L147.2 (CTS10228, CTS2180 അല്ലെങ്കിൽ Y3111 എന്ന് വിളിക്കപ്പെടുന്ന) സബ്ക്ലേഡുമായി യോജിക്കുന്ന I2a1b യുടെ ഭാഗമാണ്, ഇത് 5,600 വർഷങ്ങൾക്ക് മുമ്പ് (യമനാ കാലഘട്ടത്തിനും ട്രിപ്പിലിയൻ സ്റ്റെപ്പിലേക്കുള്ള വികാസത്തിനും തൊട്ടുമുമ്പ്) ഉടലെടുത്തതാണെന്ന് കരുതപ്പെടുന്നു, പക്ഷേ TMRCA ഉള്ളത് 2,300 മാത്രമാണ്. Yfull അനുസരിച്ച്, L147.2 നെഗറ്റീവായ I2a1b-L621 വ്യക്തികളുടെ ന്യൂനപക്ഷം കിഴക്കൻ പോളണ്ട്, ബെലാറസ്, പടിഞ്ഞാറൻ ഉക്രെയ്ൻ എന്നിവിടങ്ങളിൽ കാണപ്പെടുന്നു, ഇത് ചാൽക്കോലിത്തിക്ക് മുതൽ ഈ വംശം നിലനിന്നിരുന്നത് ഇവിടെയാണെന്ന് സൂചിപ്പിക്കുന്നു.I2a1b-L147.2 ഉപവിഭാഗം 1900 വർഷങ്ങൾക്ക് മുമ്പ് നിന്ന് വളരെ വേഗത്തിൽ വികസിച്ചു, ഇത് സ്ലാവിക് എത്നോജെനിസിസിൻ്റെ സമയവുമായി പൊരുത്തപ്പെടുന്നു, ജനസംഖ്യയുടെ തോതിൽ സ്വാധീനം ചെലുത്താൻ ഒരു മനുഷ്യന് മതിയായ പുരുഷ പിൻഗാമികൾ ഉണ്ടാകുന്നതിന് കുറച്ച് നൂറ്റാണ്ടുകൾ എടുക്കും. ഈ I2-L147.2 പൂർവ്വികൻ വളർന്നുവരുന്ന ആദ്യകാല സ്ലാവിക് ജനസംഖ്യയിൽ അത്തരമൊരു സ്വാധീനം ചെലുത്തും, 2,300 വർഷങ്ങൾക്ക് മുമ്പ് ഇപ്പോഴും ചെറുതായിരുന്നു, എന്നാൽ കുതിച്ചുയരുന്നു. കിഴക്കൻ ജർമ്മനിയിലും പോളണ്ടിലും താമസിക്കുന്ന ജർമ്മനിക് ഗോത്രങ്ങൾ, ഗോഥുകൾ, വാൻഡലുകൾ, ബർഗുണ്ടിയൻ എന്നിവരെപ്പോലെ റോമൻ സാമ്രാജ്യം ആക്രമിച്ചതിനുശേഷം, കൂടുതൽ കിഴക്ക് താമസിക്കുന്ന സ്ലാവുകൾ ശൂന്യത നികത്തി. 476-ൽ പാശ്ചാത്യ റോമൻ സാമ്രാജ്യത്തിൻ്റെ തകർച്ചയെത്തുടർന്ന്, സ്ലാവുകൾ ദിനാറിക് ആൽപ്സിലും ബാൽക്കണിലും നീങ്ങി. 9-ആം നൂറ്റാണ്ടോടെ, തുർക്കിക് സംസാരിക്കുന്ന ബൾഗറുകളുടെ നിയന്ത്രണത്തിലുള്ള കിഴക്കൻ ബാൽക്കൺ ഒഴികെയുള്ള എല്ലാ ആധുനിക സ്ലാവിക് സംസാരിക്കുന്ന പ്രദേശങ്ങളും സ്ലാവുകൾ കൈവശപ്പെടുത്തി. ഇപ്പോൾ വടക്കൻ സ്ലാവിക് രാജ്യങ്ങളിൽ I2a-L621 ൻ്റെ 9% (പോളണ്ട്, ചെക്ക് റിപ്പബ്ലിക്) നും 21% (ഉക്രെയ്ൻ) നും ഇടയിലുണ്ട്, അതേസമയം തെക്കൻ സ്ലാവുകളിൽ 20% (ബൾഗേറിയ), 50% (ബോസ്നിയ) ഉണ്ട്. തെക്ക് I2a-Din ൻ്റെ ഉയർന്ന ശതമാനം ഒരുപക്ഷേ മറ്റൊരു സ്ഥാപക പ്രഭാവം മൂലമാകാം, കാരണം തെക്കൻ സ്ലാവുകൾ പടിഞ്ഞാറൻ ഉക്രെയ്നിൽ നിന്നാണ് ഉത്ഭവിച്ചത്, അവിടെ I2a യുടെയും R1aയുടെയും അനുപാതം കൂടുതലായിരുന്നു. ഫലത്തിൽ എല്ലാ ദിനാറിക് I2a-യും L147.2 ശാഖയ്ക്ക് കീഴിലാണ്, ഭൂരിഭാഗവും 1,800 വർഷങ്ങൾക്ക് മുമ്പ് ജീവിച്ചിരുന്ന ഒരു സാധാരണ പാട്രിലീനിയർ പൂർവ്വികനിൽ നിന്ന് വന്ന S17250 റാംഫിക്കേഷനിലാണ്. ഹാപ്ലോഗ് ഗ്രൂപ്പ് I2a1b-L161.1ജനിതക വംശാവലി സർക്കിളുകളിൽ സാധാരണയായി അറിയപ്പെടുന്നത് I2-M423-ഐൽസ്, L161.1 പടിഞ്ഞാറൻ അയർലൻഡിലും (5-10%), സ്കോട്ടിഷ് ഹൈലാൻഡ്സിലും (1-5%) ഏറ്റവും ഉയർന്ന ആവൃത്തിയിൽ കാണപ്പെടുന്നു, എന്നാൽ ലാത്വിയ മുതൽ മധ്യ, പടിഞ്ഞാറൻ യൂറോപ്പിലുടനീളം കുറഞ്ഞ ആവൃത്തിയിലും (> 1%) കാണപ്പെടുന്നു, ലിത്വാനിയയും ബെലാറസും ബ്രിട്ടീഷ് ദ്വീപുകളിലേക്കും സ്കാൻഡിനേവിയ മുതൽ വടക്കുപടിഞ്ഞാറൻ സ്പെയിൻ വരെയും. അൽബേനിയ, വടക്കൻ ഗ്രീസ്, ബൾഗേറിയ, റൊമാനിയ എന്നിവിടങ്ങളിലും ഇത് കണ്ടെത്തിയിട്ടുണ്ട്. അറിയപ്പെടുന്ന ഏറ്റവും പഴയ I2a1b-L161.1 വ്യക്തികൾ മെസോലിത്തിക് ലക്സംബർഗിൽ നിന്നുള്ള 8,000 വർഷം പഴക്കമുള്ള ലോഷ്ബർ മനുഷ്യനും തെക്കൻ സ്വീഡനിലെ മൊട്ടാലയിൽ നിന്നുള്ള 7,800 വയസ്സുള്ള മനുഷ്യനുമാണ്. I2a1b-L161.1 ഒരുപക്ഷേ ഹിമാനിയുടെ അവസാനത്തിലും ഹിമയുഗാനന്തര കാലഘട്ടത്തിലും യൂറോപ്പിൻ്റെ ഭൂരിഭാഗം പ്രദേശങ്ങളിലും ചിതറിക്കിടന്നിരിക്കാം, കൂടാതെ മറ്റ് എല്ലാ മധ്യശിലായുഗ വംശങ്ങളെയും പോലെ നിയോലിത്തിക്ക് സമൂഹത്തെ സമന്വയിപ്പിച്ചിരിക്കാനാണ് സാധ്യത. I2a1b-L161.1 മെഗാലിത്തിക് കാലഘട്ടത്തിൽ ബ്രിട്ടീഷ് ദ്വീപുകളിലെ പ്രധാന നിയോലിത്തിക്ക് വംശാവലികളിൽ ഒന്നായിരുന്നു, അതിനാൽ സ്റ്റോൺഹെഞ്ച്, നോത്ത്, ന്യൂഗ്രേഞ്ച്, മറ്റ് മഹത്തായ മെഗാലിത്തിക് സൈറ്റുകൾ എന്നിവ നിർമ്മിച്ചവരിൽ ഒരാളും ഉൾപ്പെടുമായിരുന്നു. ഇന്നത്തെ അതിൻ്റെ കുറഞ്ഞ ആവൃത്തിയും യൂറോപ്പിൻ്റെ വടക്ക്-പടിഞ്ഞാറ്, തെക്ക്-കിഴക്കൻ പ്രാന്തപ്രദേശങ്ങളിലേക്കുള്ള അതിൻ്റെ ഒതുക്കവും കഴിഞ്ഞ 5,000 വർഷത്തിനിടയിലെ ഇന്തോ-യൂറോപ്യൻ കുടിയേറ്റങ്ങളുടെ നിരവധി തരംഗങ്ങളുടെ ഫലമാണ്. പ്രോട്ടോ-സെൽറ്റിക് ഹാപ്ലോഗ് ഗ്രൂപ്പ് R1b-L21 ഇപ്പോൾ അയർലണ്ടിൽ എത്തിയിരിക്കുന്നത് ഏകദേശം 2000 BCE-ലാണ് (കാസിഡി et al. 2015 കാണുക), R1b ആദ്യമായി മധ്യ യൂറോപ്പിൽ എത്തി ഏതാനും നൂറ്റാണ്ടുകൾക്ക് ശേഷമാണ്. പരിവർത്തനം അതിവേഗമായിരുന്നു, ഏതാനും നൂറ്റാണ്ടുകൾക്കുള്ളിൽ ബ്രിട്ടീഷ് ദ്വീപുകളിലെ നിയോലിത്തിക്ക് ജനസംഖ്യയെ R1b ഗോത്രങ്ങൾ കീഴടക്കുകയും അവരെ കൂടുതൽ പടിഞ്ഞാറോട്ടും വടക്കോട്ടും തള്ളിവിടുകയും ചെയ്തു, അവിടെ L161.1 ഇന്നും നിലനിൽക്കുന്നു. ഹാപ്ലോഗ് ഗ്രൂപ്പുകൾ I2-L880, I2-L1286, I2-L1294ഈ മൈനർ സബ്ക്ലേഡുകൾ പ്രധാനമായും വടക്ക്-പടിഞ്ഞാറൻ യൂറോപ്പിൽ താഴ്ന്ന ആവൃത്തിയിലാണ് കാണപ്പെടുന്നത്. I2a2 പോലെ (താഴെ കാണുക) ഈ വംശങ്ങൾ ഒരുപക്ഷേ മധ്യ, വടക്ക്-പടിഞ്ഞാറൻ യൂറോപ്യൻ വേട്ടക്കാരുടെ പിൻഗാമികളായിരിക്കാം. L880ജർമ്മനി, ലക്സംബർഗ്, ഫ്രാൻസ് എന്നിവിടങ്ങളിൽ വളരെ അപൂർവമായ ഉപവിഭാഗമാണ്. L1286പാലിയോലിത്തിക്ക് കാലഘട്ടത്തിൻ്റെ അവസാനത്തിലാണ് ഉത്ഭവിച്ചത്. എന്നിരുന്നാലും, 99% ആധുനിക വാഹകരും L233 ഉപവിഭാഗത്തിൽ പെട്ടവരും 2,000 വർഷങ്ങൾക്ക് മുമ്പ് ഒരു പൊതു പാട്രിലീനിയർ പൂർവ്വികരും പങ്കിടുന്നു. ബ്രിട്ടീഷ് ദ്വീപുകളിൽ L233 ഏറ്റവും സാധാരണമാണ്, എന്നാൽ നെതർലാൻഡ്സ്, സ്കാൻഡിനേവിയ, ജർമ്മനി, ഫ്രാൻസ്, പോളണ്ട് എന്നിവിടങ്ങളിൽ ഇടയ്ക്കിടെ കണ്ടുമുട്ടുന്നു. L1294ഇത് വളരെ അപൂർവമാണ്, ജർമ്മനി, ഫ്രാൻസ്, ബ്രിട്ടൻ എന്നിവിടങ്ങളിൽ മാത്രമേ ഇത് കണ്ടെത്തിയിട്ടുള്ളൂ. ഹാപ്ലോഗ് ഗ്രൂപ്പ് I2a2 (P214)I2a2 (S33/M436/P214, P216/S30, P217/S23, P218/S32, L35/S150, L37/S153, L181) 2005 വരെ I1c എന്നും 2010 വരെ I2b എന്നും അറിയപ്പെട്ടിരുന്നു. പ്രീ-സെൽറ്റോ-ജർമ്മനിക്വടക്കുപടിഞ്ഞാറൻ യൂറോപ്പിലെ ആളുകൾ, ഉദാഹരണത്തിന് മെഗാലിത്ത് നിർമ്മാതാക്കൾ(5000-1200 ബിസിഇ). ഇതിൻ്റെ പ്രായം 21,000 മുതൽ 13,000 വർഷം വരെ പഴക്കമുള്ളതായി കണക്കാക്കപ്പെടുന്നു, ഇത് എപ്പിപാലിയോലിത്തിക്ക് കാലഘട്ടവുമായി യോജിക്കുന്നു. I2a2 യൂറോപ്പിൻ്റെ ഭൂരിഭാഗം പ്രദേശങ്ങളിലും കാണപ്പെടുന്നു, നവീന ശിലായുഗ കർഷകരുടെ വരവിന് മുമ്പ് ഭൂഖണ്ഡത്തിലുടനീളം വ്യാപിച്ചതായി തോന്നുന്നു. നിരവധി മെസോലിത്തിക് I2a2 സാമ്പിളുകൾ ഇന്നുവരെ തിരിച്ചറിഞ്ഞിട്ടുണ്ട്, കൂടുതലും Mathieson et al. (2017) ഇതിൽ തെക്കൻ ജർമ്മനിയിൽ നിന്നുള്ള വ്യക്തികളും (സി. 7200 BCE മുതൽ M223), സെർബിയയ്ക്കും റൊമാനിയയ്ക്കും ഇടയിലുള്ള ഇരുമ്പ് ഗേറ്റുകൾ (സി. 6200 BCE മുതൽ Z161), ലാത്വിയ (സി. 5500 മുതൽ CTS10057), തെക്കുകിഴക്കൻ ഉക്രെയ്ൻ (ബി.സി.ഇ. 5400 മുതൽ L699) എന്നിവരും ഉൾപ്പെടുന്നു. C. 5200 BCE മുതൽ L701 എന്നിവയും). കൂടാതെ, സബ്ക്ലേഡ് I2c1 (L1251)അടുത്തിടെ തിരിച്ചറിഞ്ഞിട്ടുണ്ട്. ഇത് കൂടുതലും ജർമ്മനിയിലും ഇംഗ്ലണ്ടിലും അവയുടെ പ്രാന്തപ്രദേശങ്ങളിലും (അയർലൻഡ്, നോർവേ, ഫ്രാൻസ്, ഇറ്റലി, പോളണ്ട്) കാണപ്പെടുന്നു. ഈ ഉപവിഭാഗത്തിന് ഏകദേശം 6000 വർഷം പഴക്കമുണ്ടെന്ന് കരുതപ്പെടുന്നു, ഇത് നവീന ശിലായുഗത്തിൻ്റെ അവസാനത്തിൽ സ്ഥാപിക്കുന്നു. ലീനിയർ പോട്ടറി സംസ്കാരത്തിൽ നിന്ന് സമീപ കിഴക്കൻ കർഷകരുമായി സമ്പർക്കം പുലർത്തിയ ശേഷം കൃഷി സ്വീകരിച്ച ജർമ്മനിയിൽ നിന്നുള്ള വേട്ടക്കാരുമായി ഇത് ബന്ധപ്പെട്ടിരിക്കാം. പ്രശസ്ത വ്യക്തികൾI2a1b-L621 ബ്രാഞ്ച് (സ്ലാവിക്)I2a2a-Z161 ബ്രാഞ്ച് (ജർമ്മനിക്)ആൻഡ്രൂ ജോൺസൺ(1808-1875), യുണൈറ്റഡ് സ്റ്റേറ്റ്സിൻ്റെ 17-ാമത് പ്രസിഡൻ്റും 16-ാമത് വൈസ് പ്രസിഡൻ്റും ഹാപ്ലോഗ് ഗ്രൂപ്പിലെ I2a2a (മുൻ I2b1) അംഗമായി തിരിച്ചറിഞ്ഞു. പത്രത്തിൽ ആദ്യം ഹാപ്ലോഗ് ഗ്രൂപ്പ് I2a1 (മ്യൂട്ടേഷൻ P37.2) പരിഗണിക്കാം. സ്ലാവുകൾ (ഏതാണ്ട് സ്ലാവുകൾ മാത്രം!) അതിൻ്റെ ബ്രാഞ്ച് I2a1b (മ്യൂട്ടേഷനുകൾ L178/S32, M423), അല്ലെങ്കിൽ കൂടുതൽ കൃത്യമായി പറഞ്ഞാൽ, ഉപശാഖ I2a1b3 (മ്യൂട്ടേഷൻ L621/S392), അതിൻ്റെ മകൾ I2a1b3a (L147.2) എന്നിവയാണ്. കൂടുതൽ കൃത്യമായി പറയാൻ ഇപ്പോഴും ബുദ്ധിമുട്ടാണ്, കാരണം L621/S392, L147.2 മ്യൂട്ടേഷനുകളുടെ സാന്നിധ്യത്തിനായി വളരെ കുറച്ച് വ്യക്തികളെ പരീക്ഷിച്ചിട്ടുണ്ട്. സ്ലാവുകൾക്ക് പുറമേ, അവരിൽ ജർമ്മനി, ഇറ്റലി, ഗ്രേറ്റ് ബ്രിട്ടൻ എന്നിവയുടെ ഒറ്റപ്പെട്ട പ്രതിനിധികളുണ്ട്. Y ക്രോമസോമിലെ 58 ദശലക്ഷം ന്യൂക്ലിയോടൈഡുകളിൽ, സ്ലാവിക് ഉപശാഖയ്ക്ക് മാത്രമുള്ള, പിന്നീടുള്ള ഒരു മ്യൂട്ടേഷൻ കണ്ടുപിടിക്കാൻ സാധ്യതയുണ്ട്. ഹാപ്ലോഗ് ഗ്രൂപ്പ് I2a1 ൻ്റെ സ്ലാവിക് ഉപശാഖയുടെ വിതരണം ബൈനറി മാർക്കറുകളല്ല, മറിച്ച് അതിൻ്റെ സ്വഭാവ സവിശേഷതകളാൽ കണ്ടെത്താനാകും. I2a1b എന്ന ഉപബ്രാഞ്ചിൽ അവ വളരെ നിർദ്ദിഷ്ടമാണ്, ഉയർന്ന വിശ്വാസ്യതയുള്ള അതിൻ്റെ പ്രതിനിധികളെ തിരിച്ചറിയാൻ ഒരാളെ അനുവദിക്കുന്നു. ഡിഎൻഎ വംശാവലിയിൽ താൽപ്പര്യമുള്ളവർക്ക് വാണിജ്യ പരിശോധന നൽകുന്ന ഫാമിലിട്രീഡിഎൻഎയുടെ (എഫ്ടിഡിഎൻഎ) ഡാറ്റാബേസിൽ അത്തരം പരിശോധനയ്ക്കിടെ തിരിച്ചറിഞ്ഞ എല്ലാ ഹാപ്ലോടൈപ്പുകളും അടങ്ങിയിരിക്കുന്നു. ഞങ്ങൾക്ക് താൽപ്പര്യമുള്ള ഉപശാഖയെ അവിടെ "ഡൈനാറിക് ക്ലസ്റ്റർ" എന്ന് നിർവചിച്ചിരിക്കുന്നു. ഇത് രണ്ട് ശാഖകളായി വിഭജിക്കുന്നു - തെക്കൻ, യഥാർത്ഥത്തിൽ ദിനാറിക്, വടക്കൻ, എല്ലാ സ്ലാവിക് ജനസംഖ്യയിലും പ്രതിനിധീകരിക്കുന്നു (ഇതിനെ "കാർപാത്തിയൻ" എന്ന് വിളിക്കുന്നത് കൂടുതൽ ശരിയാണ്). ഈ ഡാറ്റാബേസ് ഉപയോഗിച്ച് വ്യത്യസ്ത പോപ്പുലേഷനുകളിലെ രണ്ട് ശാഖകളുടെയും ആവൃത്തി നിർണ്ണയിക്കുന്നത് അസാധ്യമാണ്, കാരണം അതിൽ അവതരിപ്പിച്ച സാമ്പിളുകൾ ക്രമരഹിതമായ അടിസ്ഥാനത്തിലാണ് രൂപപ്പെട്ടത് (ഡിഎൻഎ ടെസ്റ്റ് ഓർഡർ ചെയ്യാനുള്ള ആഗ്രഹത്തിൻ്റെയും അവസരത്തിൻ്റെയും യാദൃശ്ചികം), വലുപ്പത്തിൽ വലിയ വ്യത്യാസമുള്ളതും പ്രതിനിധികളല്ല. ജനസംഖ്യയുടെ മൊത്തത്തിൽ. ഫോറൻസിക് ആവശ്യങ്ങൾക്കായി സൃഷ്ടിച്ച് പരിപാലിക്കുന്ന വൈ-ഹാപ്ലോടൈപ്പുകളുടെ (YHRD) അന്താരാഷ്ട്ര റഫറൻസ് ഡാറ്റാബേസിലും ബെലാറസ് റിപ്പബ്ലിക്കിലെ ജനസംഖ്യയുടെ ഡിഎൻഎ മാർക്കറുകളുടെ സമാന-ഉദ്ദേശ്യ റഫറൻസ് ഡാറ്റാബേസിലും രീതിശാസ്ത്രപരമായി ശരിയായ സാമ്പിളുകൾ കണ്ടെത്താൻ കഴിയും. ബെലാറസ് റിപ്പബ്ലിക്കിലെ നീതിന്യായ മന്ത്രാലയത്തിൻ്റെ ഫോറൻസിക് എക്സ്പെർട്ടൈസ് ആൻഡ് ക്രിമിനലിസ്റ്റിക്സ് സെൻ്റർ സൃഷ്ടിച്ചത് [ബെലാറസിലെ Y-STR ലോക്കിയുടെ ഹാപ്ലോടൈപ്പുകളുടെ ആവൃത്തിയുടെ റഫറൻസ് ഡാറ്റാബേസ്: http://dnkexpertiza.org/index1.html ]. YHRD-യിൽ ഉൾപ്പെടാത്ത ചില സാമ്പിളുകളുടെ ഹാപ്ലോടൈപ്പുകൾ ശാസ്ത്രീയ പ്രസിദ്ധീകരണങ്ങളിൽ കാണാം. ഈ സാമ്പിളുകളെല്ലാം FTDNA യുമായി താരതമ്യപ്പെടുത്തുമ്പോൾ താരതമ്യേന കുറഞ്ഞ എണ്ണം ലോക്കികളാണ് പ്രതിനിധീകരിക്കുന്നത്: വാണിജ്യ പരിശോധനകളിൽ 37, 67, 111 ലോക്കുകൾ എന്നിവയും ഉപയോഗിക്കുന്നു, ഫോറൻസിക് പരിശോധനകൾ 17-ലോകസ് സെറ്റ് ഉപയോഗിക്കുന്നു, ഇത് Y-Filer സ്റ്റാൻഡേർഡ് എന്നറിയപ്പെടുന്നു. . എന്നാൽ ഈ സെറ്റ് പോലും ദിനാറിക് ക്ലസ്റ്ററിൻ്റെ സവിശേഷതയായ ഹാപ്ലോടൈപ്പുകൾ ആത്മവിശ്വാസത്തോടെ തിരിച്ചറിയുന്നത് സാധ്യമാക്കുന്നു. ഈ ക്ലസ്റ്ററിനായുള്ള മോഡൽ ഹാപ്ലോടൈപ്പിൽ നിന്നുള്ള താരതമ്യേന ചെറിയ ജനിതക ദൂരമാണ് ഇവയുടെ സവിശേഷത, ഇത് Y-Filer ഫോർമാറ്റിൽ പട്ടിക 1 ൽ കാണിച്ചിരിക്കുന്നു. പട്ടിക 1. ഹാപ്ലോഗ് ഗ്രൂപ്പ് I2a1-ൻ്റെ ദിനാറിക് ക്ലസ്റ്ററിൻ്റെ മോഡൽ ഹാപ്ലോടൈപ്പ്
ഈ പ്രൊഫൈലിൽ നിന്ന് മൊത്തത്തിൽ 9-10 ഘട്ടങ്ങളിൽ കൂടുതൽ വ്യത്യാസമുള്ള ഹാപ്ലോടൈപ്പുകൾ (ആവർത്തനങ്ങളുടെ എണ്ണത്തിലെ മ്യൂട്ടേഷനുകൾ) മിക്ക കേസുകളിലും ഞങ്ങൾക്ക് താൽപ്പര്യമുള്ള I2a1 ശാഖയിൽ പെടുന്നു. അവരുടെ നിർണ്ണയത്തിലെ സാധ്യമായ പിശക് 1-2% കവിയരുത്, ബൈനറി മാർക്കറുകൾ ഉപയോഗിച്ച് ഒരേസമയം സാമ്പിളുകൾ പരീക്ഷിച്ച സന്ദർഭങ്ങളിൽ (അയ്യോ, കുറച്ച്) ഇത് സ്ഥിരീകരിക്കുന്നു. ദിനാറിക് ക്ലസ്റ്ററിൻ്റെ തെക്കൻ ബ്രാഞ്ച് 67-ഉം അതിലും കൂടുതലായി 111 ലോക്കികളോടും കൂടി വേറിട്ടുനിൽക്കുന്നു, എന്നാൽ 17-ലോക്കസ് Y-ഫയലർ സെറ്റിൽ, DYS-448 ലോക്കസിലെ ആവർത്തനങ്ങളുടെ എണ്ണം കുറയുന്നു എന്നതാണ് ഒരേയൊരു സവിശേഷത: 19 ( ഇടയ്ക്കിടെ പോലും 18) 20 എന്നതിനുപകരം. തീർച്ചയായും, രണ്ട് ഉപക്ലസ്റ്ററുകൾ തമ്മിലുള്ള വ്യത്യാസം ഒരു ആദ്യ ഏകദേശമെന്ന നിലയിൽ മാത്രമേ സാധ്യമാകൂ, എന്നാൽ ഇത്രയും ഏകദേശ കണക്കെടുത്താൽ പോലും മൊത്തത്തിലുള്ള ചിത്രം വ്യക്തമാണ്. Y-Filer ഫോർമാറ്റിൻ്റെ എല്ലാ പ്രസിദ്ധീകരിച്ച സാമ്പിളുകൾക്കുമായി ഈ ക്ലസ്റ്ററിൽ ഉൾപ്പെടുന്ന ഹാപ്ലോടൈപ്പുകളുടെ ആവൃത്തികൾ പട്ടിക 2 ൽ അവതരിപ്പിച്ചിരിക്കുന്നു. പട്ടിക 2. ദിനാറിക് ക്ലസ്റ്ററിൻ്റെ ഹാപ്ലോടൈപ്പുകളുടെ ആവൃത്തി
ഹാപ്ലോഗ് ഗ്രൂപ്പ് I2a1 ൻ്റെ ദിനാറിക് ക്ലസ്റ്ററിന് ബാൽക്കൻ-കാർപാത്തിയൻ മേഖലയിലും പടിഞ്ഞാറൻ, കിഴക്കൻ സ്ലാവുകൾക്കിടയിലും - ലൂക്കാ-റേക്കോവറ്റ്സ് സംസ്കാരത്തിൻ്റെ മേഖലയിലും പരമാവധി ഉണ്ടെന്ന് പട്ടിക കാണിക്കുന്നു. പോളണ്ടിലും റഷ്യയിലും അതിൻ്റെ ആവൃത്തി കുറയുന്നു, ലിത്വാനിയയിലും ജർമ്മനിയിലും ഇത് സ്ലാവിക് മിശ്രിതത്താൽ നന്നായി വിശദീകരിക്കാൻ കഴിയുന്ന ഒരു തലത്തിലാണ്. ഇതിനർത്ഥം ലുസാഷ്യൻ-പോമറേനിയൻ പ്രദേശത്ത് നിന്ന് പ്രോട്ടോ-സ്ലാവിക് ഭാഷ കൊണ്ടുവന്ന ബസ്താർനിക്കും വെനെറ്റിക്കും ഇപ്പോഴും ഈ ക്ലസ്റ്റർ ഇല്ലായിരുന്നു - അതുപോലെ തന്നെ സ്ലാവുകളുമായുള്ള തീവ്രമായ സമ്പർക്കം ആരംഭിക്കുന്നതിന് മുമ്പ് പുരാതന ബാൾട്ടുകൾക്കും. ഓട്ടോക്ത്തോണസ് ജനസംഖ്യയുടെ ഒരു പ്രധാന ഗ്രൂപ്പിൻ്റെ സ്വാംശീകരണത്തിൻ്റെ ഫലമായി ഇത് ആധുനിക സ്ലാവുകളുടെ ജീൻ പൂളിൽ പ്രവേശിച്ചു. എല്ലാ നോൺ-സ്ലാവിക് ജനങ്ങളിലും, ഹാപ്ലോഗ് ഗ്രൂപ്പ് I2a1 ന് റൊമാനിയക്കാർക്കിടയിൽ മാത്രമേ ഉയർന്ന ആവൃത്തിയുള്ളൂ, അവർ ആദ്യകാല സ്ലാവുകളുടെ (പ്രാഗ്, ഇപോട്ടെസ്റ്റി-ചുറേലിയൻ സംസ്കാരങ്ങളുടെ വാഹകർ) വ്ലാച്ചുകളുമായുള്ള മിശ്രിതത്തിൻ്റെ ഫലമാണ് - ത്രേസിയൻ്റെ പിൻഗാമികൾ. റോമൻ സാമ്രാജ്യത്തിൻ്റെ കാലഘട്ടത്തിൽ ജനസംഖ്യ റോമൻവൽക്കരിക്കപ്പെട്ടു. മാത്രമല്ല, റൊമാനിയക്കാർക്കിടയിലും സ്ലാവിക് വംശീയ ഗ്രൂപ്പിൻ്റെ (എൽവോവ് മേഖല, തെക്കൻ ബെലാറസ്) രൂപീകരണ മേഖലയിലും ആവൃത്തി ഏകദേശം തുല്യമാണ്. തൽഫലമായി, വ്ലാച്ചുകൾക്കിടയിൽ ഇത് തുടക്കത്തിൽ ആദ്യകാല സ്ലാവുകളേക്കാൾ കുറവായിരുന്നില്ല. തെക്കൻ ഉപക്ലസ്റ്ററിൻ്റെ വിതരണം സൂചകമാണ്. ഇത് പൊതുവെ സ്ലാവിക് ജനസംഖ്യയുടെ സവിശേഷതയല്ല. കിഴക്കൻ സ്ലാവിക്, പോളിഷ് സാമ്പിളുകളിൽ കാണപ്പെടുന്ന DYS448=19 ഉള്ള കുറച്ച് ഹാപ്ലോടൈപ്പുകൾ പോലും ശേഷിക്കുന്ന സ്ഥലങ്ങളുടെ മോഡൽ മൂല്യങ്ങളിൽ നിന്ന് വളരെ അകലെയാണ്, മാത്രമല്ല സ്വതന്ത്രമായ മ്യൂട്ടേഷനുകളെ പ്രതിഫലിപ്പിക്കുകയും ചെയ്യുന്നു. അതേ സമയം, സൗത്ത് സ്ലാവിക് ഹാപ്ലോടൈപ്പുകൾക്ക് പലപ്പോഴും മോഡൽ ഒന്നിനോട് വളരെ അടുത്തുള്ള ഒരു പ്രൊഫൈൽ ഉണ്ട്. അവരുടെ വാഹകരിൽ ഭൂരിഭാഗവും DYS448 ലോക്കസിൽ 19 ആവർത്തനങ്ങൾ നടത്തിയ ഒരു വ്യക്തിയുടെ പിൻഗാമികളാണ്. ഈ വ്യക്തി മുഴുവൻ ദിനാറിക് ക്ലസ്റ്ററിൻ്റെ പൂർവ്വികനേക്കാൾ പിന്നീട് ജീവിച്ചു, പക്ഷേ അധികം അല്ല - പൊതുവേ, തെക്കൻ ഉപക്ലസ്റ്ററിലെ ഹാപ്ലോടൈപ്പുകളുടെ വ്യാപനം വളരെ വലുതാണ്, പൂർവ്വിക ഹാപ്ലോടൈപ്പിൽ നിന്ന് 6-7 പടികളുടെ ജനിതക ദൂരം അതിൽ അസാധാരണമല്ല. 20 ആവർത്തനങ്ങളുള്ള പൂർവ്വിക ക്ലസ്റ്ററും 19 ആവർത്തനങ്ങളുള്ള മകൾ ഉപക്ലസ്റ്ററും, വെങ്കലയുഗത്തിൻ്റെ അവസാനത്തോടെ - ഇരുമ്പ് യുഗത്തിൻ്റെ ആരംഭം, പ്രധാനമായും കാർപാത്തിയൻസിന് തെക്ക് വിതരണം ചെയ്യപ്പെട്ടു, കൂടാതെ, മകൾ ക്ലസ്റ്റർ തെക്കൻ ഭാഗത്ത് ഉയർന്നുവന്നു. ശ്രേണിയുടെ ഭാഗവും സ്ലാവുകൾ പ്രത്യക്ഷപ്പെടുന്നതിന് മുമ്പ് മുഴുവൻ കാർപാത്തിയൻ തടത്തിലേക്കും വ്യാപിക്കാൻ സമയമില്ലായിരുന്നു. എഡി 271-ൽ ഡാസിയ ഒഴിപ്പിച്ചതിനുശേഷം. ഇ. പ്രായോഗികമായി ഡാന്യൂബ് കടക്കാത്ത ബൈസൻ്റൈൻ പ്രജകളുടെ മാത്രം സ്വഭാവമായിരുന്നു അത്. ഇതിനിടയിൽ, DYS448=20 ഉള്ള പഴയതും കൂടുതൽ വിപുലവുമായ ഒരു ഉപവിഭാഗം കാർപാത്തിയൻസിന് വടക്ക് തുളച്ചു കയറി, അവിടെ അത് വളർന്നുവരുന്ന സ്ലാവുകളുടെ ഭാഗമായി. എപ്പോൾ, എവിടെയാണ് സ്ലാവുകളുടെ പൂർവ്വികർ "ദിനാറിക്" ഹാപ്ലോടൈപ്പുകളുടെ ഉയർന്ന അനുപാതമുള്ള ഒരു ഗ്രൂപ്പിനെ ഉൾപ്പെടുത്തിയത്? ഇത് രണ്ടാം നൂറ്റാണ്ടിന് മുമ്പ് സംഭവിക്കാൻ പാടില്ലായിരുന്നു. ബി.സി. - സരുബിൻസി സംസ്കാരത്തിൻ്റെ രൂപീകരണ സമയം. എന്നാൽ ഇത് 7-8 നൂറ്റാണ്ടുകളുടെ തുടക്കത്തിന് ശേഷം സംഭവിക്കില്ലായിരുന്നു, വടക്കോട്ട് മടങ്ങിവരുന്ന കുടിയേറ്റ പ്രക്രിയയിൽ ലൂക്കാ-റൈക്കോവറ്റ്സ് സംസ്കാരം ഉയർന്നുവന്നു. റഷ്യക്കാർക്കും ധ്രുവങ്ങൾക്കുമിടയിൽ I2a1 ൻ്റെ ആധുനിക ആവൃത്തി വിശദീകരിക്കാൻ മറ്റൊരു മാർഗവുമില്ല, ഈ ഹാപ്ലോഗ് ഗ്രൂപ്പ് തുടക്കത്തിൽ ഇല്ലാതിരുന്ന പ്രദേശങ്ങളിൽ രൂപപ്പെട്ടു. ഇവിടെ രണ്ട് സാഹചര്യങ്ങൾ സാധ്യമാണ്. I2a1 ൻ്റെ വാഹകർ സരുബിൻസി സംസ്കാരത്തിൻ്റെ രൂപീകരണത്തിൽ പങ്കെടുത്ത പ്രാദേശിക സംസ്കാരങ്ങളിലൊന്നിൻ്റെ പ്രതിനിധികളാകാം അല്ലെങ്കിൽ അന്തരിച്ച സരുബിൻസി ഗ്രൂപ്പുകളിൽ ഒന്നായിരിക്കാം. മിലോഗ്രാഡ്-ന്യൂഴ്സ്, ബോറിസ്ഫെനിറ്റുകൾ-സിഥിയൻസ്, അല്ലെങ്കിൽ യുഖ്നോവൈറ്റ്സ്-മെലാഞ്ച്ലെൻസ് എന്നിവയെക്കുറിച്ച് നമുക്ക് സംസാരിക്കാം. രണ്ടാമത്തെ സാഹചര്യത്തിൽ, ഉക്രെയ്നിലെ ചെർനിയാഖോവ് സംസ്കാരത്തിൻ്റെ അവശിഷ്ടങ്ങൾ അല്ലെങ്കിൽ കാർപാത്തിയൻ തടത്തിലെയും മധ്യ ഡാന്യൂബിലെയും റോമലൈസ്ഡ് ത്രേസിയൻസിൻ്റെ അവശിഷ്ടങ്ങൾ സ്വാംശീകരിക്കുന്നതിനിടയിൽ, വളർന്നുവരുന്ന സ്ലാവുകളുടെ കുടിയേറ്റ പ്രക്രിയയിൽ ജീൻ പൂളിൻ്റെ ഈ ഘടകം ഇതിനകം തന്നെ നേടിയെടുത്തു. വ്യത്യസ്ത ഗവേഷകർക്കിടയിൽ അതിൻ്റെ പങ്കിനെക്കുറിച്ചുള്ള വിലയിരുത്തലുകൾ വ്യത്യസ്തമാണെങ്കിലും, സറൂബിൻസി സംസ്കാരത്തിൽ (പ്രത്യേകിച്ച് അതിൻ്റെ അപ്പർ ഡൈനിപ്പർ വേരിയൻ്റിൽ) മിലോഗ്ഗ്രാഡ് അടിവസ്ത്രത്തിൻ്റെ സാന്നിധ്യം വളരെ സാധ്യതയുണ്ട്. മിഡിൽ ഡൈനിപ്പർ വേരിയൻ്റിൽ ലേറ്റ് സിഥിയൻ അടിവസ്ത്രം വ്യക്തമായി കാണാം. കീവ് സംസ്കാരത്തിൻ്റെ രൂപീകരണത്തിൽ നിർണ്ണായക പങ്ക് വഹിച്ചിട്ടുണ്ടെങ്കിലും, പോച്ചെപ്പ്, കർത്തമിഷെവ്, ഗ്രിനി-അബിദ്നിയ തരം - പരേതനായ സരുബിൻസി ഗ്രൂപ്പുകളുടെ രൂപീകരണത്തിൽ ഒരു പ്രധാന യുഖ്നോവ്സ്കി ഘടകം പങ്കെടുത്തു. എന്നാൽ ഈ സാഹചര്യം വടക്കൻ, തെക്കൻ ഉപക്ലസ്റ്ററുകളുടെ ഹാപ്ലോടൈപ്പുകളുടെ ഉയർന്ന സാമ്യത്തെ മോശമായി വിശദീകരിക്കുന്നു. യുഖ്നോവ് സംസ്കാരത്തിൻ്റെ ഉത്ഭവത്തെക്കുറിച്ച് അറിയപ്പെടുന്നത് ബാൽക്കൻ-കാർപാത്തിയൻ പ്രദേശത്തെ സംസ്കാരങ്ങൾക്ക് പൊതുവായ ഒരു ഘടകത്തിൻ്റെ സാന്നിധ്യത്തെക്കുറിച്ച് സംസാരിക്കാൻ ഞങ്ങളെ അനുവദിക്കുന്നില്ല. മിലോഗ്രാഡ്, ഫോറസ്റ്റ്-സ്റ്റെപ്പ് സിഥിയൻ സംസ്കാരങ്ങളിൽ, അത്തരമൊരു ഘടകം ഒരു പ്രത്യേക തെക്കുപടിഞ്ഞാറൻ പ്രേരണയായിരിക്കാം, ഇത് ബിസി ഒന്നാം സഹസ്രാബ്ദത്തിൻ്റെ തുടക്കത്തിൽ ചെർണോൾസ് സംസ്കാരത്തിൻ്റെ രൂപീകരണ സമയത്ത് രേഖപ്പെടുത്തിയിട്ടുണ്ട്. ഇ. സ്റ്റാമ്പ് ചെയ്തതും ഫ്ലൂട്ട് ചെയ്തതുമായ സെറാമിക്സിൻ്റെ പ്രോട്ടോ-ത്രേസിയൻ സ്രഷ്ടാക്കളുമായി ബന്ധപ്പെട്ട ഒരു ജനസംഖ്യയ്ക്ക് "ഡൈനാറിക്" ഹാപ്ലോടൈപ്പുകളെ ഉക്രേനിയൻ ഫോറസ്റ്റ്-സ്റ്റെപ്പുകളിലേക്ക് കൊണ്ടുവരാൻ കഴിയുമായിരുന്നു, തുടർന്ന്, തുടർച്ചയായി അവ സിഥിയൻമാർക്കും മിലോഗ്രാഡിയൻമാർക്കും കൈമാറാൻ കഴിയുമായിരുന്നു. എന്നാൽ ഈ ഹാപ്ലോടൈപ്പുകളുടെ വാഹകർ നിരവധി നൂറ്റാണ്ടുകളായി പ്രധാന ത്രേസിയൻ ലോകത്ത് നിന്ന് ഒറ്റപ്പെട്ട് നിലനിന്നിരിക്കണം, ശ്രദ്ധേയമായ നിരവധി നിർദ്ദിഷ്ട മ്യൂട്ടേഷനുകൾ ശേഖരിച്ചു. അതേസമയം, ബെലാറസിൻ്റെയും റഷ്യയുടെയും പ്രദേശത്ത് I2a1 ഹാപ്ലോടൈപ്പുകളിൽ പ്രത്യേക പ്രത്യേകതകളൊന്നുമില്ല. രണ്ടാമത്തേതിൽ, കൂടുതൽ സാധ്യതയുള്ള സാഹചര്യത്തിൽ, നോർത്ത് ദിനാറിക് ഹാപ്ലോടൈപ്പുകളുടെ ദാതാക്കൾ കാർപാത്തിയൻ മൗണ്ട് സംസ്കാരമാണ്, വികാസ പ്രക്രിയയിൽ പ്രാഗ് സംസ്കാരം ആഗിരണം ചെയ്യുന്നു (ഇത് കാർപ്പ് ഗോത്രവുമായി തിരിച്ചറിയപ്പെടുന്നു), ഡാക്കോ-റോമൻ്റെ ശേഷിക്കുന്ന ഗ്രൂപ്പുകൾ. കാർപാത്തിയൻ തടത്തിലെ ജനസംഖ്യ. അവാർ ഖഗാനേറ്റിനുള്ളിൽ സ്ലാവുകൾ അവരുമായി ഇടകലർന്നപ്പോൾ, വടക്കൻ ഉപവിഭാഗത്തിൻ്റെ ആവൃത്തി ഏകദേശം 20% ആയി നിശ്ചയിച്ചു. തുടർന്ന് അവർ തങ്ങളുടെ ഹാപ്ലോടൈപ്പുകൾ ഭാഗികമായി വനമേഖലയിലേക്ക് കൊണ്ടുപോയി, അവിടെ പ്രാദേശിക ജനസംഖ്യയുമായി ഇടകലർന്നതിനാൽ ആവൃത്തി കുറഞ്ഞു. കൂടുതൽ തെക്ക്, ബൈസൻ്റൈൻ സാമ്രാജ്യത്തിലേക്ക് പോയവർ, സൗത്ത് ദിനാറിക് ഹാപ്ലോടൈപ്പുകളുടെ ഉയർന്ന സാന്ദ്രതയുള്ള ഓട്ടോചോണുകളെ അവിടെ കണ്ടുമുട്ടി. ക്രൊയേഷ്യക്കാരും സെർബുകളും ഭാഗികമായി മാസിഡോണിയക്കാരും പൊതുവെ I2a1 ൻ്റെ ഉയർന്ന ആവൃത്തിക്ക് കടപ്പെട്ടിരിക്കുന്നത് ഈ അടിവസ്ത്രത്തിലാണ്, ഇത് പൊതു സ്ലാവിക് ലെവലിനെ ഗണ്യമായി കവിയുന്നു. ഡാന്യൂബിന് തെക്ക് സ്ഥിരതാമസമാക്കിയ സ്ലാവുകൾ മടക്ക കുടിയേറ്റത്തിൽ മിക്കവാറും പങ്കെടുത്തില്ല, അതിനാലാണ് വനമേഖലയിൽ കുറച്ച് സൗത്ത് ദിനാറിക് ഹാപ്ലോടൈപ്പുകൾ ഉണ്ടായിരുന്നത്. ഡാന്യൂബിൻ്റെ വടക്ക് ഭാഗത്തേക്ക് ഈ ഹാപ്ലോടൈപ്പുകളെ കൂട്ടത്തോടെ കൊണ്ടുവന്നത് ഓസ്ട്രിയക്കാരുടെ സ്ലാവിക് പൂർവ്വികരും അതുപോലെ റൊമാനിയക്കാരുടെ പൂർവ്വികരായ വ്ലാച്ചുകളും മാത്രമാണ്. Y-ക്രോമസോമൽ ഹാപ്ലോഗ് ഗ്രൂപ്പ് I2a യുടെ ഉത്ഭവം, അതിൻ്റെ പരിണാമം, ഈ ഹാപ്ലോഗ് ഗ്രൂപ്പിൻ്റെ വാഹകരുടെ ജനസംഖ്യയുടെ മൈഗ്രേഷൻ റൂട്ടുകൾ എന്നിവ 5 വർഷമായി എൻ്റെ പ്രത്യേക പഠനത്തിന് വിഷയമാണ്, മനുഷ്യ ജനസംഖ്യയെക്കുറിച്ചുള്ള സജീവമായ പഠനത്തിൽ എനിക്ക് താൽപ്പര്യമുണ്ടായ സമയം മുതൽ. ജനിതകശാസ്ത്രം. ഈ ഹാപ്ലോഗ് ഗ്രൂപ്പിലുള്ള എൻ്റെ താൽപ്പര്യത്തിൻ്റെ കാരണം വളരെ ലളിതമാണ് - എൻ്റെ Y-ക്രോമസോമിൻ്റെ (Y-STR + Y-snips) ഒരു ജനിതക വിശകലനം കാണിക്കുന്നത്, ഫൈലോജെനെറ്റിക് വർഗ്ഗീകരണത്തിൻ്റെ വീക്ഷണകോണിൽ, ഇത് ഇതിൻ്റെ ഒരു ശാഖയിൽ പെടുന്നു എന്നാണ്. ഹാപ്ലോഗ് ഗ്രൂപ്പ്, ഈ നിന്ദ്യമായ ജൈവ വസ്തുത കാരണം, അതിൽ ഞാൻ ഈ ഹാപ്ലോഗ് ഗ്രൂപ്പിൻ്റെ പുരാതന പ്രതിനിധികളിൽ നിന്നുള്ള നേരിട്ടുള്ള പുരുഷ നിരയിൽ നിന്നാണ് വരുന്നത്. ഈ ഹാപ്ലോഗ് ഗ്രൂപ്പിനെ കുറിച്ചുള്ള പഠനത്തിൽ ഞാൻ ഏർപ്പെടാനുള്ള ഒരു പ്രധാന കാരണം I2 ൻ്റെയും മുഴുവൻ ഹാപ്ലോഗ് ഗ്രൂപ്പിൻ്റെയും നിർഭാഗ്യകരമായ അറിവില്ലായ്മയാണ്. ഈ ഹാപ്ലോഗ് ഗ്രൂപ്പിനെക്കുറിച്ചുള്ള ഏറ്റവും പുതിയ പ്രത്യേക പഠനങ്ങൾ 2004, 2006, 2007-ൽ പ്രസിദ്ധീകരിച്ചുവെന്ന് പറഞ്ഞാൽ മതി - റൂട്ട്സി , സിരി തുടങ്ങിയവർ.(2004), Rootsi, S. (2006), Peter Underhill et al (2007). ബാക്കിയുള്ള കൃതികൾ, പ്രത്യേകിച്ച് ബാൽക്കണിലെയും മുൻ യുഗോസ്ലാവിയയിലെയും ജനസംഖ്യയുടെ Y-ക്രോമസോം വൈവിധ്യത്തെ സംബന്ധിച്ചുള്ളവ, പ്രധാനമായും മുകളിൽ പറഞ്ഞ മൂന്ന് കൃതികളിൽ പറഞ്ഞിരിക്കുന്ന നിഗമനങ്ങൾ, അവരുടേതായ ഒന്നും ചേർക്കാതെ തന്നെ ആവർത്തിക്കുന്നു (Marjanovic, Damir; et al.2005, Peričić , മരിജാന തുടങ്ങിയവർ. 2005; റെബാല കെ. തുടങ്ങിയവർ.(2007). അതിനാൽ, എസ്റ്റോണിയൻ പോപ്ജെനെറ്റിസിസ്റ്റ് ഗവേഷകനായ റൂട്ട്സിയുടെ സൃഷ്ടിയാണ് ഹാപ്ലോഗ് ഗ്രൂപ്പ് I ൻ്റെ ഉത്ഭവത്തെക്കുറിച്ചുള്ള നിരവധി അടിസ്ഥാന സിദ്ധാന്തങ്ങൾ നിർണ്ണയിച്ചത്, അവ പിന്നീട് ശാസ്ത്രീയവും ജനപ്രിയവുമായ ശാസ്ത്ര (വിക്കിപീഡിയ) ഉറവിടങ്ങളിൽ ഏകീകരിക്കപ്പെട്ടു. നിർഭാഗ്യവശാൽ, ഈ അനുമാനങ്ങളിൽ പലതും തെറ്റായിരുന്നു. ഔദ്യോഗിക യാഥാസ്ഥിതിക ജനസംഖ്യാ ജനിതകശാസ്ത്രത്തിൽ പൊതുവായി അംഗീകരിക്കപ്പെട്ടിട്ടുള്ള I2 നെക്കുറിച്ചുള്ള വിക്കിപീഡിയ ലേഖനത്തിൽ അടങ്ങിയിരിക്കുന്ന ചില വ്യവസ്ഥകൾ ഞങ്ങൾ ചുവടെ അവതരിപ്പിക്കും, കൂടാതെ ലേഖനം വിലയിരുത്തുമ്പോൾ, മുകളിലുള്ള ചില വ്യവസ്ഥകൾ പിന്നീട് പുനർവിചിന്തനം ചെയ്യുകയും അമച്വർമാരുടെ ജനിതകശാസ്ത്രത്തിന് സമീപമുള്ള സർക്കിളുകളിൽ വികസിപ്പിക്കുകയും ചെയ്തു.
ഹാപ്ലോഗ് ഗ്രൂപ്പും ഭാഷയും, ഹാപ്ലോഗ് ഗ്രൂപ്പും പുരാവസ്തു സംസ്കാരവും (ഈ കണക്ഷനുകൾ ഒരു പ്രത്യേക പഠനം അർഹിക്കുന്നു) തമ്മിലുള്ള ബന്ധത്തെക്കുറിച്ചുള്ള ചോദ്യം മാറ്റിനിർത്തിയാൽ, ബാൽക്കൻ പൂർവ്വിക മാതൃരാജ്യത്തിന് അനുകൂലമായ റൂട്ട്സിയുടെ പ്രധാന വാദം ദിനാറിക്കിലെ ഈ ഹാപ്ലോഗ് ഗ്രൂപ്പിൻ്റെ പ്രധാന സാന്നിധ്യമാണെന്ന് നമുക്ക് കാണാം. ആൽപ്സ്/ബാൾക്കൻസ് മേഖല. ഏറ്റവും ലളിതവും വിലകുറഞ്ഞതും, ഒരു പാഴ്സിമോണി വീക്ഷണകോണിൽ നിന്ന്, അത്തരം സംഖ്യകൾക്കുള്ള വിശദീകരണം, യൂറോപ്പിലെ അവസാന ഹിമയുഗം (20 ആയിരം വർഷങ്ങൾക്ക് മുമ്പ്), അതായത്, ബാൽക്കൻ ജനസംഖ്യയുടെ ജീൻ പൂളിൻ്റെ ഒരു ഭാഗത്തിൻ്റെ തുടർച്ചയെ അനുമാനിക്കുക എന്നതാണ്. ബാൽക്കൻ ഗ്ലേഷ്യൽ അഭയകേന്ദ്രത്തിൻ്റെ കാലം മുതൽ. നിങ്ങൾക്കറിയാവുന്നതുപോലെ, Y-ക്രോമസോം ഹാപ്ലോഗ് ഗ്രൂപ്പുകളുടെ ഭൂമിശാസ്ത്രപരമായ ഉത്ഭവം ഇനിപ്പറയുന്ന അനുഭവപരമായ മാനദണ്ഡമനുസരിച്ച് സ്ഥാപിക്കാവുന്നതാണ്: ഒരു നിശ്ചിത ഹാപ്ലോഗ് ഗ്രൂപ്പ് ഉണ്ടായ സ്ഥലത്ത് അല്ലെങ്കിൽ ജനസംഖ്യയിൽ, അതിൻ്റെ ആവൃത്തിയും STR വിതരണവും (അല്ലെങ്കിൽ STR വേരിയബിളിറ്റിയുടെ പ്രായം)മറ്റ് ജനസംഖ്യയുമായി താരതമ്യം ചെയ്യുമ്പോൾ പരമാവധി (സെൻഗുപ്ത et al., 2006). ഹാപ്ലോഗ് ഗ്രൂപ്പ് I ൻ്റെ ഉദാഹരണം ഉപയോഗിച്ച് കൂടുതൽ വിശദമായി പരിശോധിക്കുമ്പോൾ, ഈ ലളിതവും യുക്തിസഹമായി തോന്നുന്നതുമായ സ്കീം, പുതിയ ഡാറ്റയുടെ വെളിച്ചത്തിൽ വിമർശനത്തിന് എതിരല്ല. എന്നിരുന്നാലും, ജനിതക ശാസ്ത്രജ്ഞർക്ക് ഹാപ്ലോഗ് ഗ്രൂപ്പ് I യുടെ ഘടനയെക്കുറിച്ച് പൊതുവായ ധാരണ മാത്രമുണ്ടായിരുന്ന കാലത്താണ് റൂട്ട്സിയുടെ ലേഖനങ്ങളുടെ പശ്ചാത്തലം സജ്ജീകരിച്ചത് എന്നത് ഓർമ്മിക്കേണ്ടതാണ്. സ്പെഷ്യലൈസ്ഡ് പോപ്പുലേഷൻ ജനിതകശാസ്ത്രജ്ഞരുടെ തുടർന്നുള്ള ഗവേഷണവും ഹാപ്ലോഗ് ഗ്രൂപ്പ് I-ൻ്റെ സമർപ്പിത അമേച്വർ പഠനവും. വാണിജ്യ പദ്ധതികൾ ഡിഎൻഎ വംശാവലി എന്ന് വിളിക്കപ്പെടുന്ന വ്യക്തിഗത അമേച്വർ ഗവേഷകർ (കെന്നത്ത് നോർഡ്വെഡ്റ്റ്, ബെർണി കുള്ളൻ, സ്റ്റീവ് ട്രാങ്സ്രുഡ്, വില്യം ഹാർട്ട്ലി, യെമെ ഡ്രോസ്റ്റ്, ആരോൺ ജെ. ഹിൽ, പീറ്റർ ഹ്രെക്ഡാകിയൻ, ഹൊവൻ സിമോണിയൻ, ബോബ് മെയ്, വാഡിം വെറെനിച്, ബിൽ മാർഡ് സെയ്ക, ലോമർ, ലോമർ മുതലായവ) ഹാപ്ലോഗ് ഗ്രൂപ്പിൻ്റെ ശാഖകളുടെ-ഉപക്ലേഡുകളുടെ ഫൈലോജെനെറ്റിക് ബന്ധങ്ങളും ഘടനകളും കൂടുതൽ നന്നായി മനസ്സിലാക്കാൻ അനുവദിച്ചു, കൂടാതെ ഹാപ്ലോഗ് ഗ്രൂപ്പിൻ്റെ വ്യക്തിഗത ശാഖകൾ-ഉപക്ലേഡുകൾക്ക് കാലക്രമവും ഭൂമിശാസ്ത്രപരവുമായ റഫറൻസ് നൽകുന്നു. ഹാപ്ലോഗ് ഗ്രൂപ്പ് I2a യുടെ ഉത്ഭവത്തെയും പരിണാമത്തെയും കുറിച്ചുള്ള നമ്മുടെ അറിവിൻ്റെ നിലവിലെ അവസ്ഥയുടെ ഒരു ഹ്രസ്വ സ്നാപ്പ്ഷോട്ട് മാത്രമായി ഈ പഠനം പരിമിതപ്പെടുത്തും. ഹാപ്ലോഗ് ഗ്രൂപ്പ് I2a യുടെ ഉത്ഭവവും മൈഗ്രേഷൻ റൂട്ടുകളും. അവലോകനത്തിൻ്റെ ശിഥിലമായ സ്വഭാവം കാരണം, യൂറോപ്പിൽ ഞാൻ എങ്ങനെ, എപ്പോൾ ഹാപ്ലോഗ് ഗ്രൂപ്പ് പ്രത്യക്ഷപ്പെട്ടു എന്നതിനെക്കുറിച്ചുള്ള തുല്യ പ്രാധാന്യമുള്ള ചോദ്യങ്ങളും അതുപോലെ തന്നെ ബേസൽ ബ്രാഞ്ച് I ഹാപ്ലോഗ് ഗ്രൂപ്പുകളായി I1, I2 എന്നിവയായി വിഭജിച്ച സമയവും സ്ഥലവും സംബന്ധിച്ച ചോദ്യങ്ങളും ഞങ്ങൾ സ്പർശിക്കില്ല. നിലവിൽ, ഈ സാഹചര്യത്തിൻ്റെ നിരവധി, തുല്യ സാധ്യതയുള്ള പതിപ്പുകളിലൊന്നിന് അനുകൂലമായി ഞങ്ങൾക്ക് മതിയായ തെളിവുകൾ ഇല്ല, അതിനാൽ മധ്യ പാലിയോലിത്തിക്ക് യൂറോപ്പിലെ നിവാസികളുടെ ഡിഎൻഎയെക്കുറിച്ചുള്ള വരാനിരിക്കുന്ന പഠനത്തിൻ്റെ പ്രതീക്ഷയിൽ ഞങ്ങൾ ഈ ചോദ്യങ്ങൾ ഒഴിവാക്കും. പകരം, ഈ ഗ്രൂപ്പിൻ്റെ മറ്റ് ശാഖകൾ മാറ്റിവെച്ച് ഹാപ്ലോഗ് ഗ്രൂപ്പ് I2a-യുടെ ഫൈലോജെനെറ്റിക് ('കുടുംബ വൃക്ഷം') ട്രീ അവതരിപ്പിച്ചുകൊണ്ട് ഞങ്ങൾ വിവരണം ആരംഭിക്കും. ഹാപ്ലോഗ് ഗ്രൂപ്പ് I ൻ്റെ സാധാരണ ലളിതമായ ഡയഗ്രമുകൾക്ക് പകരം, നോർഡ്വെഡ്റ്റിൻ്റെ പ്രോസസ്സിംഗിൽ (പതിപ്പ് 2013) വൃക്ഷത്തിൻ്റെ കാനോനിക്കൽ പതിപ്പ് ഞങ്ങൾ വാഗ്ദാനം ചെയ്യുന്നു. ഉപക്ലേഡുകളുടെയും ഹാപ്ലോടൈപ്പുകളുടെ ഇൻട്രാക്ലേഡ് ക്ലസ്റ്ററുകളുടെയും ശാഖകളുടെ ആപേക്ഷിക ടോപ്പോളജിക്കൽ ക്രമീകരണത്തിൻ്റെ ദൃശ്യപരമായ പ്രാതിനിധ്യമാണ് ഇതിൻ്റെ പ്രധാന നേട്ടം. കൂടാതെ, കെൻ നോർഡ്വെഡ്റ്റ് ക്രോസ്-ഡിസ്പെർഷൻ രീതി ഉപയോഗിച്ച് കണക്കാക്കിയ ശാഖകളുടെ വ്യതിചലനത്തിൻ്റെ (വ്യതിചലനം) ഇൻ്റർക്ലേഡ് സമയം കണക്കിലെടുത്താണ് മരത്തിന് താഴെയുള്ള സ്കെയിൽ നിർമ്മിച്ചിരിക്കുന്നത്. അവസാനമായി, ഫൈലോജെനെറ്റിക് ട്രീയുടെ ഗ്രാഫിക്കൽ പ്രാതിനിധ്യം ചരിത്രാതീത യൂറോപ്പിൻ്റെ ക്രോണോടോപ്പിലെ ഹാപ്ലോഗ് ഗ്രൂപ്പ് I ൻ്റെ വാഹകരുടെ ചലനത്തിൻ്റെ സംയോജിത പുനർനിർമ്മാണത്താൽ പൂർത്തീകരിക്കപ്പെടുന്നു. ഹാപ്ലോഗ് ഗ്രൂപ്പ് I2a യുടെ വിഭജനത്തിൻ്റെ കാലഗണന ഇപ്രകാരമാണ് (ഞങ്ങളുടെ പതിപ്പ്): ഏകദേശം 14,000-15,000 വർഷങ്ങൾക്ക് മുമ്പ് I2a L460/PF3647/S2381ഒരു ശാഖ പ്രത്യക്ഷപ്പെട്ടു I2a2-L35/PF3862/S150, L37/PF6900/S153, L181, M436/P214/PF3856/S33, P216/PF3855/S30, P217/PF3854/S23/S23, P321, അത് ഏതാണ്ട് ഉടനടി വിഭജിക്കപ്പെടുന്നു I2a2a-L34/PF3857/S151, L36/S152, L59, L368, L622, M223, P219/PF3859/S24, ഇതിന് സമാന്തരമായി, ഹാപ്ലോഗ് ഗ്രൂപ്പായ I2a-ൽ നിന്ന് ഒരു ഉപശാഖ ഉയർന്നുവന്നു I2a1-P37.2 (15,000-10,000 വർഷം മുമ്പ്). വിപുലമായ വിശകലനത്തിൽ ഒരു പ്രത്യേക ഹാപ്ലോഗ് ഗ്രൂപ്പ് I2a1 തിരിച്ചറിഞ്ഞുവെന്നത് ശ്രദ്ധേയമാണ് -F (ഐഎസ്ഒജിജി ട്രീയിൽ ഉൾപ്പെടുത്തിയിട്ടില്ലാത്ത L1294 L1297 L1298 സ്നിപ്പുകൾ തിരിച്ചറിയുന്നു). Nordtvedt കണക്കാക്കിയ ഇൻ്റർക്ലേഡ് യുഗങ്ങൾ അനുസരിച്ച്, ഈ ക്ലസ്റ്റർ I2a1a ഗ്രൂപ്പിനേക്കാൾ പിന്നീട് ഉയർന്നുവന്നു, എന്നാൽ I2a1b ഗ്രൂപ്പിനേക്കാൾ മുമ്പാണ് (അതായത്, രണ്ട് ശാഖകൾക്കിടയിലുള്ള മധ്യത്തിൽ). Nordtvedt അനുസരിച്ച്, I2a1 ൻ്റെ അവസാന പൊതു പൂർവ്വികൻ-F, I2a1a എന്നിവ നമ്മുടെ കാലത്തിന് 20,000 വർഷങ്ങൾക്ക് മുമ്പ് ജീവിച്ചിരിക്കില്ല. I2a1*-F ൻ്റെ പ്രതിനിധികൾ പ്രധാനമായും ആധുനിക പടിഞ്ഞാറൻ ഫ്രാൻസിൻ്റെ പ്രദേശത്താണ് താമസിക്കുന്നത് എന്നത് സവിശേഷതയാണ്. കൂടാതെ, I2a1c യുടെ പ്രതിനിധികളും പടിഞ്ഞാറൻ യൂറോപ്പിൽ താമസിക്കുന്നു. നിന്ന് I2a1-P37.2ബിസി 20,000 നും 15,000 നും ഇടയിൽപെട്ടി അവൻ്റെ സമയം വേറിട്ടു നിന്നുI2a1aL158/PF4073/S433, L159.1/S169.1, M26/PF4056((അവസാന പൊതു പൂർവ്വികൻ്റെ ജീവിതകാലം - - 8,000-6,000) കൂടാതെI2a1b-M423 L178/S328, M423(അവസാനത്തെ പൊതു പൂർവ്വികൻ്റെ ജീവിതകാലം ഇപ്പോഴുള്ളതിന് 14,000 വർഷത്തിൽ കുറയാത്തതാണ്) I2a1b-M423 L178/S328ഇൻ്റർക്ലേഡ് അക്കൌണ്ട് അനുസരിച്ച്, ഏകദേശം 13,500 വർഷങ്ങൾക്ക് മുമ്പ് പൂർവ്വിക രേഖകൾ വേർതിരിച്ചു.I2a1b3, I2a1b2 - (Dinaric+Disles) കൂടാതെ ദ്വീപുകൾ. കൗതുകകരമെന്നു പറയട്ടെ, ദ്വീപുകളുടെ ഇന്നത്തെ കാലഘട്ടത്തിന് ഏകദേശം 6600 വർഷങ്ങൾക്ക് മുമ്പുള്ള അവസാന പൊതു പൂർവ്വികൻ്റെ ആയുസ്സ് ഏകദേശം 8000 വർഷമാണെന്ന് ഞങ്ങൾ കണക്കാക്കുന്നു.
അതിലും കൂടുതൽ വൈകിയുള്ള ഇൻട്രാക്ലേഡ് ഡിവിഷനുകൾ (atകൃത്യം 8 ആയിരം-6 ആയിരം വർഷങ്ങൾക്ക് മുമ്പ്) ca യുടെ ആഗോള തണുപ്പുമായി നന്നായി ബന്ധപ്പെട്ടിരിക്കുന്നു. 6200 ബിസി, യൂറോപ്പിലെ ജനസംഖ്യ വ്യക്തമായി കുറഞ്ഞപ്പോൾ വിദഗ്ധർ രേഖപ്പെടുത്തിയ നിയോലിത്തിക്ക് ജനസംഖ്യാ പ്രതിസന്ധിയിലേക്ക് നയിച്ചു.അതേ സമയം വടക്കൻ യൂറോപ്പിൽഎൻ്റെ സമയം (ഏകദേശം 6500 -6200 BC) ഭൂമിശാസ്ത്രപരമായി വേർതിരിക്കുന്ന (ഫൈലോജെനറ്റിക് ആയി നേരത്തെ വേർതിരിക്കപ്പെട്ട) ജനസംഖ്യ I2a1b2-I2a1b3 എന്നിവ സംഭവിച്ചു. , അതുപോലെ തന്നെ I2a2-M284-ൻ്റെ ഭാഗമാണ് - ബാക്കിയുള്ള I2a2 ഉപക്ലേഡുകളിൽ നിന്ന് ഐൽസ് സബ്ക്ലേഡ്.ഡോഗർലാൻഡിലെ വെള്ളപ്പൊക്കത്തെക്കുറിച്ചാണ് നമ്മൾ സംസാരിക്കുന്നത്. ഉപയോഗിച്ച് നിർവചിച്ചിരിക്കുന്നത്ആധുനിക കടൽത്തീരത്തിൻ്റെ രൂപരേഖ ഉൾപ്പെടെയുള്ള തെളിവുകൾ കാണിക്കുന്നത്, അവസാനത്തെ ഹിമാനിയുടെ സമയത്ത് വടക്കൻ കടലിനും ഇംഗ്ലീഷ് ചാനലിനുമിടയിലുള്ള നീർത്തടങ്ങൾ കിഴക്കൻ ആംഗ്ലിയയിൽ നിന്ന് തെക്ക് കിഴക്ക് ഹോക്ക് വാൻ ഹോളണ്ട് വരെ, പാസ് ഡി കാലായിസ് വഴിയല്ല, തേംസ് നദികൾ ഒഴുകിയിരുന്നെന്നും മ്യൂസും ഷെൽഡും റൈനും ചേർന്ന് ഇംഗ്ലീഷ് ചാനലിലൂടെ വിശാലമായ ഒരു നദിയുടെ രൂപത്തിൽ ഒഴുകി, ഒടുവിൽ അത് അറ്റ്ലാൻ്റിക് സമുദ്രത്തിലേക്ക് ഒഴുകി.[ഏകദേശം 8000 BC. ഇ. ഭൂപ്രദേശത്തിൻ്റെ വടക്കൻ തീരം, ഡോഗ്ഗെലഗൂണുകൾ, ചതുപ്പുകൾ, വേലിയേറ്റ തീരങ്ങൾ, ബീച്ചുകൾ എന്നിവയുള്ള ഒരു തീരപ്രദേശമായിരുന്നു rland. ഒരുപക്ഷേ മധ്യശിലായുഗത്തിൽ ഈ പ്രദേശം വേട്ടയാടൽ, കോഴി, മത്സ്യബന്ധനം എന്നിവയുടെ കാര്യത്തിൽ യൂറോപ്പിലെ ഏറ്റവും സമ്പന്നമായിരുന്നു. ഹിമാനിയുടെ ഉരുകലിൻ്റെ ഫലമായി സമുദ്രനിരപ്പിലുണ്ടായ ക്രമാനുഗതമായ വർദ്ധനവ് (അവസാന ഹിമാനിയുടെ അവസാനം) വടക്കൻ കടലിനാൽ ഡോഗർലാൻഡ് വെള്ളപ്പൊക്കത്തിലേക്ക് നയിച്ചുവെന്ന് അനുമാനിക്കപ്പെടുന്നു, ഇത് 6500-ഓടെ യൂറോപ്യൻ ഭൂഖണ്ഡത്തിൽ നിന്ന് ചരിത്രാതീത ബ്രിട്ടനെ ക്രമേണ വെട്ടിമാറ്റി. ബി.സി. ഇ. നിലവിൽ ഡോഗർ ബാങ്ക് എന്നറിയപ്പെടുന്ന ഭാവി സാൻഡ്ബാങ്കിൻ്റെ സൈറ്റിൽ, ഈ സിദ്ധാന്തമനുസരിച്ച്, കുറഞ്ഞത് 5000 ബിസി വരെ ദ്വീപ് നിലനിന്നിരുന്നു. ഇ. 8,200 വർഷങ്ങൾക്ക് മുമ്പ് (ബിസി 6,200) ഒരു സുനാമി മൂലം ഭൂരിഭാഗം പ്രദേശങ്ങളും വെള്ളത്തിനടിയിലായി എന്നതാണ് അടുത്തിടെ മുന്നോട്ട് വച്ച ഒരു ബദൽ സിദ്ധാന്തം. ചരിത്രത്തിലെ ഏറ്റവും വലിയ ഒന്ന്മണ്ണിടിച്ചിൽ - തീരത്ത് നിന്ന് വെള്ളത്തിനടിയിലുള്ള മണ്ണിൻ്റെ വിസ്തീർണ്ണംനോർവേ സ്റ്റുറെഗ്ഗ എന്നറിയപ്പെടുന്നു . ഈ സിദ്ധാന്തമനുസരിച്ച്, മണ്ണിടിച്ചിൽ മൂലമുണ്ടായ സുനാമി പ്രദേശത്തെ മെസോലിത്തിക് ജനസംഖ്യയ്ക്ക് ഒരു വിനാശകരമായ പങ്ക് വഹിച്ചു. സുനാമിയുടെ ഫലമായി, ബ്രിട്ടൻ ഭൂഖണ്ഡത്തിൽ നിന്ന് ശാശ്വതമായി വേർപിരിഞ്ഞതായി കാണപ്പെടുന്നു, സാംസ്കാരിക വീക്ഷണത്തിൽ,മധ്യശിലായുഗം അതിൽ സ്വന്തം വഴിക്ക് പോയി. പാർശ്വഫലങ്ങളിൽ ഒന്ന്ഒരു മണ്ണിടിച്ചിൽ ഉണ്ടായിആഗോള തണുപ്പിക്കൽ 6200 ബിസി ഇ. ഉരുകിയ ഹിമാനികളിൽ നിന്നുള്ള തണുത്ത വെള്ളത്തിൻ്റെ വരവ് കാരണം. ഹാപ്ലോഗ് ഗ്രൂപ്പ് I2a യുടെ ചരിത്രാതീത കാലത്തെ പ്രതിനിധികൾ ആരായിരുന്നു? ഐ. ഹാപ്ലോഗ് ഗ്രൂപ്പ് I2a യുടെ പൂർവ്വികർ എവിടെയാണ് പ്രത്യക്ഷപ്പെട്ടത്?? വിക്കിപീഡിയയിൽ നിന്നുള്ള മേൽപ്പറഞ്ഞ ഉദ്ധരണിയിൽ നിന്ന് കാണാൻ കഴിയുന്നത് പോലെ, ചില ജനിതക വൃത്തങ്ങൾക്കിടയിൽ, തെറ്റായ അഭിപ്രായം, ഞങ്ങളുടെ അഭിപ്രായത്തിൽ, ട്രിപ്പിലിയൻസിന് സമീപമുള്ള സംസ്കാരങ്ങൾ ഉൾപ്പെടെയുള്ള ബാൽക്കൻ നിയോലിത്തിക്ക് സംസ്കാരങ്ങളുടെ സർക്കിളുമായി I2a ബന്ധപ്പെട്ടിരിക്കുന്നു. ഈ ഓപ്ഷൻ സാധ്യമാണ്, എന്നാൽ ഈ സംസ്കാരങ്ങളുടെ പ്രതിനിധികളുടെ അവശിഷ്ടങ്ങളുടെ ഡിഎൻഎയേക്കാൾ പഴയ വൈ-ക്രോമസോമുകളുടെ വിശകലനത്തിലൂടെ അന്തിമ പതിപ്പ് നിർണ്ണയിക്കണം. പക്ഷേ, ട്രിപ്പിലിയൻമാരിൽ I2 ഉണ്ടായിരുന്നെങ്കിൽ, അവയെ ഒരു തദ്ദേശീയ ഘടകമായി കണക്കാക്കാനാവില്ല (ഇതിൻ്റെ കാതൽ മിഡിൽ ഈസ്റ്റിൽ നിന്നുള്ള കുടിയേറ്റക്കാരായിരുന്നു); മിക്കവാറും, അവർ ഈ സംസ്കാരത്തിൽ ഉൾപ്പെടുത്തിയിട്ടുള്ള ഒരു പ്രാദേശിക മെസോലിത്തിക്ക് മൂലകത്തിൻ്റെ അവശിഷ്ടങ്ങളെ പ്രതിനിധീകരിക്കുന്നു. പുരാതന I2a2 ൻ്റെ ആദിമ സംസ്കാരത്തെക്കുറിച്ച് നമ്മൾ സംസാരിക്കുകയാണെങ്കിൽ, അവർ മിക്കവാറും സ്വിഡേറിയൻമാരും അവരുടെ എപ്പിഗോണുകളും ആയിരുന്നു. 2011-ൽ, ഹാപ്ലോഗ് ഗ്രൂപ്പായ I2a1b-M423 (പൊതുവായി I2a) യുടെ ഉത്ഭവത്തെക്കുറിച്ച് പോളിഷ് FTDNA പ്രോജക്റ്റിൻ്റെ (ലാറി മജ്ക) അഡ്മിനിസ്ട്രേറ്ററുമായി ഒരു ചെറിയ ചർച്ച നടത്താൻ എനിക്ക് കഴിഞ്ഞു. ഇതിനുമുമ്പ്, മൈക്കിന് ഈ ഹാപ്ലോഗ് ഗ്രൂപ്പിൻ്റെ ഉത്ഭവത്തെക്കുറിച്ച് താരതമ്യേന വലിയ താൽപ്പര്യമില്ലായിരുന്നു (എന്നിരുന്നാലും, എന്നെപ്പോലെ, അവനും അതിൽ ഉൾപ്പെടുന്നു). ഹാപ്ലോഗ് ഗ്രൂപ്പ് I2a1 ൻ്റെ ഉത്ഭവ സ്ഥലത്തെക്കുറിച്ച് വളരെ രസകരമായ ഒരു പതിപ്പ് അദ്ദേഹം നിർദ്ദേശിച്ചു. അദ്ദേഹത്തിൻ്റെ അഭിപ്രായത്തിൽ, ഉയർന്ന തോതിലുള്ള സംഭാവ്യതയുള്ള എല്ലാ I2a1 ൻ്റെയും (I2a1*, I2a1a, I2a1b, I2a1c) പൊതു പൂർവ്വികൻ ആൽപൈൻ മേഖലയിൽ ജീവിച്ചിരിക്കാം. ആൽപ്സ് പർവതനിരകളിൽ നിന്ന് കാർപാത്തിയൻസിലേക്ക് I2a1 വംശം കടന്നുപോകുന്നതിനുള്ള എൻ്റെ നിർദ്ദേശിച്ച വഴിയും അദ്ദേഹം അംഗീകരിച്ചു. ശരിയാണ്, അവൻ എൻ്റെ ഡേറ്റിംഗിനോട് യോജിച്ചില്ല. I2a1 ൻ്റെ പൂർവ്വിക ജനസംഖ്യ ആൽപ്സിൽ നിന്ന് കാർപാത്തിയനുകളിലേക്കുള്ള (പടിഞ്ഞാറൻ യൂറോപ്പിൽ നിന്ന് വടക്കോട്ട് പിൻവാങ്ങുന്ന ഹിമാനിയെ തുടർന്ന്) അതിൻ്റെ ചലനം ആരംഭിച്ചുവെന്ന് ഞാൻ വിശ്വസിക്കുന്നു, LGM അവസാനിച്ചതിന് തൊട്ടുപിന്നാലെ (അതായത് ബിസി 10,000 ന് ശേഷമുള്ള മധ്യശിലായുഗത്തിൽ) മൈക്ക് വിശ്വസിച്ചു. കുടിയേറ്റത്തിൻ്റെ തുടക്കക്കാരൻ സെൽറ്റുകളായിരിക്കാം (അതായത്, ഈ കുടിയേറ്റം ഇതിനകം വെങ്കലയുഗത്തിൽ സംഭവിച്ചു). പിന്നീടുള്ള ഓപ്ഷൻ, രസകരമാണെങ്കിലും, ദ്വീപ് സബ്ക്ലേഡ് I2a1b2, ദിനാറിക് സബ്ക്ലേഡ് I2a1b3 എന്നിവ വേർതിരിക്കുന്ന സമയവും സ്ഥലവും വിശദീകരിക്കുന്നില്ല, തന്മാത്രാ ഡേറ്റിംഗ് അനുസരിച്ച്, 10-8 ആയിരം വർഷങ്ങൾക്ക് മുമ്പ് ഇത് സംഭവിക്കില്ല. മൈക്കിയുടെ പതിപ്പ് പരിശോധിക്കുന്നതിന്, നിലവിലെ ഭൂമിശാസ്ത്രപരമായ വിതരണത്തെ (ഭൂമിശാസ്ത്രപരമായ) അടിസ്ഥാനമാക്കി സബ്ക്ലേഡിൻ്റെ “അടിത്തറ” യുടെ ഏറ്റവും സാധ്യതയുള്ള (പാർസിമണിയുടെ വീക്ഷണകോണിൽ നിന്ന്) സ്ഥലം കണക്കാക്കാൻ ഞങ്ങളെ അനുവദിക്കുന്ന മെസ്ക്വിറ്റ് അൽഗോരിതം ഉപയോഗിക്കാൻ ഞാൻ തീരുമാനിച്ചു. സ്പ്രെഡ്) രണ്ട് അടുത്തുള്ള സബ്ക്ലേഡുകളുടെ (ഈ സാഹചര്യത്തിൽ, ഹാപ്ലോടൈപ്പ് ടാക്സയുടെ ഭൂമിശാസ്ത്രപരമായ കോർഡിനേറ്റുകളുടെയും I2a1b ട്രീയുടെ ടോപ്പോളജിയുടെയും താരതമ്യത്തെ അടിസ്ഥാനമാക്കിയാണ് ഞങ്ങൾ I2a1b1 ദൃശ്യമാകുന്ന സ്ഥലം നിർണ്ണയിക്കുന്നത്, ഇത് ഔട്ട്ഗ്രൂപ്പ് റൂട്ട് I2a1a വഴി വേരൂന്നിയതാണ്). നിലവിൽ I2a1a സബ്ക്ലേഡിൻ്റെ ഏറ്റവും ഉയർന്ന സാന്ദ്രത സാർഡിനിയയിലാണ് എന്ന വസ്തുത ഉണ്ടായിരുന്നിട്ടും, നോർഡ്വെഡ്റ്റിൻ്റെ നന്നായി യുക്തിസഹവും അനുഭവപരമായി സ്ഥിരീകരിച്ചതുമായ അഭിപ്രായമനുസരിച്ച്, പ്രായത്തിലുള്ള ഏറ്റവും പഴയ I2a1a ക്ലസ്റ്ററുകൾ വടക്കൻ മേഖലയിലാണ്. ഐബീരിയൻ പെനിൻസുല, ആൽപ്സ്, തെക്കൻ ഫ്രാൻസിൻ്റെ ചില ഭാഗങ്ങൾ. ഈ ആവശ്യത്തിനായി, 2D-3D പ്രൊജക്ഷനിൽ, വടക്കൻ യൂറോപ്പിൻ്റെ ഭൂമിശാസ്ത്രപരമായ പ്രൊജക്ഷനിലേക്ക് മെസ്ക്വിറ്റിലെ ഫൈലോജെനെറ്റിക് ട്രീകൾ എങ്ങനെ പ്രയോഗിക്കാമെന്ന് ഞാൻ പഠിച്ചു. ഉദാഹരണത്തിന്, 687 ഹാപ്ലോടൈപ്പുകൾ I2a1b-M423 ഉള്ള ഒരു ഫൈലോജെനെറ്റിക് ട്രീയുടെ ഘടനയെ അടിസ്ഥാനമാക്കി ഞാൻ മെസ്ക്വിറ്റ് കാർട്ടോഗ്രാഫറിൽ നിർമ്മിച്ച 2 മാപ്പുകൾ, ടിഎൻടിയിലെ 20 മികച്ച (പാർസിമോണിക്) മരങ്ങളുടെ 20 സ്വതന്ത്ര പൊരുത്തങ്ങൾ കണ്ടെത്തി. ചുവന്ന നിറം പുനർനിർമ്മിച്ച ഉത്ഭവസ്ഥാനത്തെ സൂചിപ്പിക്കുന്നു - ഈ സാഹചര്യത്തിൽ ആൽപൈൻ പ്രദേശം.
അതിനാൽ, I2a1b കാരിയറുകൾക്കായി ഞങ്ങൾക്ക് ലഭിച്ച പാത ഞങ്ങൾ നിർദ്ദേശിച്ച റൂട്ടുമായി ഏകദേശം യോജിക്കുന്നു. I2a1b യും I2a1b3 യും പ്രത്യക്ഷപ്പെടുന്നതിന് ഇടയിൽ ആയിരക്കണക്കിന് വർഷങ്ങൾ കടന്നുപോയതിനാൽ, കുടിയേറ്റം എപ്പോഴാണ് സംഭവിച്ചതെന്ന് കൃത്യമായി പറയാൻ പ്രയാസമാണ്. മറ്റൊരു കാര്യം പ്രധാനമാണ്, പ്രസ്ഥാനം ആരംഭിച്ചത് ബാൽക്കണിൻ്റെ പ്രദേശത്ത് നിന്നല്ല, മറിച്ച് ഫ്രഞ്ച് അല്ലെങ്കിൽ സ്വിസ് ആൽപ്സ് മുതൽ വടക്കുകിഴക്ക് വരെയാണെന്ന് ഞങ്ങൾ കാണിച്ചു. ഈ പതിപ്പിനെ ഭൗതിക തെളിവുകൾ - പുരാതന ഡിഎൻഎ മുതലായവ പിന്തുണയ്ക്കുന്നുണ്ടോ? അക്ഷരാർത്ഥത്തിൽ വളരെ അടുത്ത കാലം വരെ, ഞങ്ങളുടെ നിർദ്ദിഷ്ട സിദ്ധാന്തം അധിക സ്ഥിരീകരണം ആവശ്യമായ ഒരു സ്റ്റാറ്റിസ്റ്റിക്കൽ ഇൻ്റർപോളേഷൻ മാത്രമായി തുടർന്നു. രണ്ടാഴ്ച മുമ്പ്, 'പ്രൊഫഷണൽ' ജനിതകശാസ്ത്രജ്ഞരുടെ കൃതി ഒടുവിൽ പ്രസിദ്ധീകരിച്ചു, ഇത് ഇന്നുവരെ പഠിച്ച പ്രാചീന സാമ്പിളുകളുടെ മിക്ക ഡിഎൻഎയും ആധുനിക മനുഷ്യ ജനസംഖ്യയുടെ ഡിഎൻഎയുമായി എങ്ങനെ ബന്ധപ്പെട്ടിരിക്കുന്നുവെന്ന് കാണിക്കുന്നു ( പുരാതന മനുഷ്യ ജീനോമുകൾ ഇന്നത്തെ യൂറോപ്യന്മാർക്ക് മൂന്ന് പൂർവ്വിക ജനസംഖ്യ നിർദ്ദേശിക്കുന്നു). കഴിഞ്ഞ വർഷങ്ങളിൽ ഞാൻ ശബ്ദമുയർത്തിയ I2a1b യുടെ ഉദ്ഭവസ്ഥാനത്തെയും കുടിയേറ്റത്തിൻ്റെ തുടക്കത്തെയും കുറിച്ചുള്ള എൻ്റെ അനുമാനത്തിന് സൃഷ്ടിയിൽ കാര്യമായ സ്ഥിരീകരണം ലഭിച്ചു. ഒന്നാമതായി, യൂണിപാരൻ്റൽ ലൈനുകളിൽ, സ്വീഡിഷ്, ലക്സംബർഗ് മെസോലിത്തിക് വേട്ടക്കാരുടെ പുരാതന ഡിഎൻഎ ജീൻ പൂൾ mtDNA ഹാപ്ലോഗ് ഗ്രൂപ്പ് യു, വൈ-ക്രോമസോമൽ ഹാപ്ലോഗ് ഗ്രൂപ്പ് I എന്നിവയുടെ പ്രതിനിധികളായി മാറി, അതിനാൽ ഞങ്ങൾ വീണ്ടും തുടക്കത്തിൽ തന്നെ R1a/R1b യുടെ അഭാവം കൈകാര്യം ചെയ്യുന്നു. മെസോലിത്തിക്ക് യൂറോപ്യന്മാർ. രണ്ടാമതായി, ഈ പഠനങ്ങൾ അത് കാണിക്കുന്നു യുക്സംബർഗ്മെസോലിത്തിക്ക് വേട്ടക്കാരൻ (ഇന്നത്തേതിന് ഏകദേശം 8000 വർഷങ്ങൾക്ക് മുമ്പ് ലക്സംബർഗിലെ ഹെഫിംഗൻ-ലോച്ച്ബർഗിലെ മെസോലിത്തിക്ക് സൈറ്റിൻ്റെ പര്യവേക്ഷണത്തിനിടെ കണ്ടെത്തിയ അവശിഷ്ടങ്ങൾ) ഹാപ്ലോഗ് ഗ്രൂപ്പിൽ പെടുന്നു. I2a1b* M423 + L178+. പട്ടികയിൽ നൽകിയിരിക്കുന്ന Y-ക്രോമസോം പോളിമോർഫിസങ്ങളെക്കുറിച്ചുള്ള ഡാറ്റ അനുസരിച്ച്, ഈ ചരിത്രാതീത ലക്സംബർഗിൻ്റെ ഹാപ്ലോഗ് ഗ്രൂപ്പ് I2a1b2, I2a1b3 എന്നിവയുമായി ബന്ധപ്പെട്ട് പൂർവ്വികരാണ്. I2a1b L178 G A 15574052 12 + മൂന്നാമതായി, ഈ പഠനങ്ങൾ കാണിക്കുന്നത് മൊട്ടാലയിലെ മെസോലിത്തിക്ക് ജനസംഖ്യയുടെ ഒരു ഭാഗവും (സ്വീഡൻ, ഓസ്റ്റ്ഗേറ്റർലാൻഡ്, ഇപ്പോഴത്തേതിന് ഏകദേശം 8000 വർഷങ്ങൾക്ക് മുമ്പ്) I2a1b*(xI2a1b1, I2a1b3) |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||






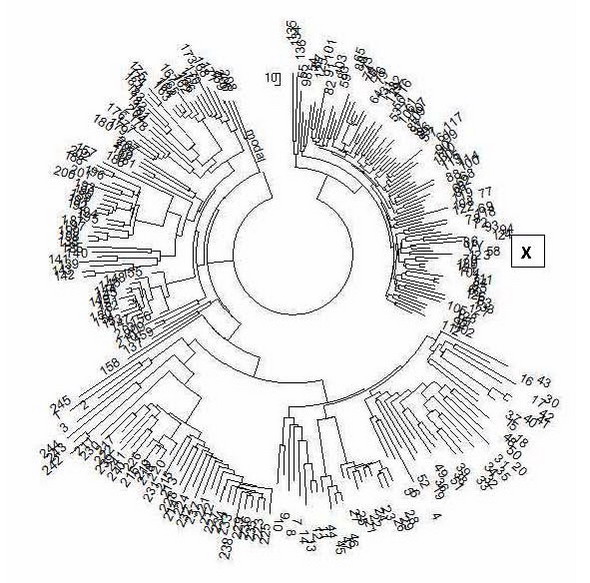













ജനപ്രിയമായത്:

പുതിയത്
- ശിക്ഷിക്കപ്പെട്ട ഒരു വ്യക്തിയുടെ സാമൂഹികവൽക്കരണത്തിൻ്റെ 14 പ്രധാന ദിശകൾ
- വ്യാസെസ്ലാവ് നാഗോവിറ്റ്സിൻ ഒരു ജോലിയും കണ്ടെത്തിയില്ല
- അലക്സി പുഷ്കോവ്: ജീവചരിത്രം, വ്യക്തിഗത ജീവിതം, കുടുംബം, ഫോട്ടോ ബാല്യം, കുടുംബം
- പെച്ചെർസ്കിലെ വിശുദ്ധ സിസോയ്, സ്കീമാമോങ്ക്, പെചെർസ്കിലെ വിശുദ്ധ വെനറബിൾ സ്കീമാമോങ്ക് ജോസാഫ്
- യേശുക്രിസ്തുവിൻ്റെ പുനരുത്ഥാനം ക്രിസ്തുവിൻ്റെ പുനരുത്ഥാനത്തിനുശേഷം സംഭവിച്ചത്
- കത്തീഡ്രൽ ഓഫ് ബ്ലെസ്ഡ് വിർജിൻ മേരി കത്തീഡ്രൽ
- ഓർത്തഡോക്സ് എൻസൈക്ലോപീഡിയ ട്രീയിലെ നാലാമത്തെ മെനയോണിൻ്റെ അർത്ഥം
- ആരോഗ്യ മന്ത്രാലയത്തിൻ്റെ ഉത്തരവ് 290n തീയതി 02
- എക്സ്പോണൻഷ്യൽ അസമത്വങ്ങൾ
- സ്ഥാപിത മാനദണ്ഡങ്ങൾ അടിസ്ഥാനമാക്കി, ജോലിഭാരത്തിൻ്റെ മാനദണ്ഡങ്ങൾ, ഹെഡ്കൗണ്ട് മാനദണ്ഡങ്ങൾ, ഡോക്ടർമാരുടെ മറ്റ് തൊഴിൽ മാനദണ്ഡങ്ങൾ എന്നിവ കണക്കാക്കുന്നു.




