Vietnes sadaļas
Redaktora izvēle:
- No valodniecības vēstures Izmantotās literatūras saraksts
- Altaja valodu saimē ietilpst
- Gruzīnu valodas un tūrisma vārdnīca
- Ir tikai strukturālās un objektu programmēšanas paradigmas Programmēšanas paradigmas un tehnoloģijas
- Armēņu alfabēts kā ķīmisko elementu periodiskās tabulas kods
- Slāvi un substrāts (fragments no pārskatītās versijas par haplogrupu I2a1)
- "Fets Afanasijs Afanasjevičs
- Ģeodēziskie darbi lineāriem objektiem
- Balmonta dzejoļa “Es nācu šajā pasaulē...
- Gatavs elektroniskais portfolio bērnudārza audzinātājai
Reklāma
| I2 haplogrupa. Slāvi un substrāts (fragments no pārskatītās versijas par haplogrupu I2a1) |
|
111 marķieru haplotipi tika ņemti par visdetalizētākajiem, un izrādījās, ka starp tiem ir 15 haplotipi ar pirmajiem 12 marķieriem, kas ir tieši tādi paši kā uzrādītajā haplotipā. Tie ir haplotipi ar numuriem 57-60, 110, 131, 132, 153-157, 660, 744, 745 saskaņā ar 968 haplogrupas I1 111 marķieru haplotipu sarakstu. Pēc tam, izmantojot profesionālu datorprogrammu (PHYLIP), no visiem 968 111 marķieru haplotipiem tika uzbūvēts koks, un tika identificēti vissaistītākie (pēc pirmajiem 12 marķieriem) haplotipi. Iegūtais koks ilustratīviem nolūkiem ir parādīts zemāk. Tas parāda labu simetriju, kas liecina, ka viss koks ir cēlies no viena kopīga senča. Kad viņš dzīvoja, arī tiks parādīts. Informācijai visa koka pamata (senču) haplotips ir šāds 13 22 14 10 13 14 11 14 11 12 11 28 – 15 8 9 8 11 23 16 20 28 12 14 15 16 – 10 10 19 21 14 14 16 20 35 37 12 10 – 11 8 15 15 8 11 10 8 9 9 12 23 25 15 10 12 12 16 8 13 25 20 13 13 11 12 11 11 12 11 – 32 12 8 17 12 24 27 19 11 12 12 13 11 9 11 11 10 12 12 31 11 13 21 16 11 10 24 15 19 11 24 17 13 15 25 12 22 18 12 14 18 9 12 11 Uzrādītais haplotips pirmajos 12 marķieros no tā atšķiras tikai ar vienu mutāciju (piektā mutācija pēc kārtas, izcelta). Kopumā visi 968 haplotipi parāda 14 891 mutāciju no pirmā 67 marķieru paneļa un 24 990 mutācijas uz visiem 111 marķieriem. Tas parāda, ka kopējais sencis dzīvoja 14891/968/0,12 = 128 → 147 parastās paaudzes (katra 25 gadi) vai 24990/968/0,198 = 130 → 149 tradicionālās paaudzes (0,12 un 0,198 ir mutācijas ātruma konstantes un 1 117 mutācijas ātruma konstantes). marķieru haplotipi, bultiņa – tabulas korekcija atkārtotām mutācijām), vai attiecīgi 3675±370 un 3725±375 gadus pirms kopējā priekšteča. Var redzēt, ka 67 un 111 marķieru haplotipi dod gandrīz identiskus rezultātus ar tikai 50 gadu atšķirību uz 3700 gadu fona.
Izrādījās, ka šie septiņi haplotipi, kas satur vienādus 12 marķierus/alēles, atrodas trīs dažādos koka zaros. Protams, ja uzrādītajam haplotipam būtu 25, 37 vai vairāk marķieri, noteikti būtu izveidojies tikai viens zars, bet tas ir tāds, kāds tas ir. Principā tas mūs īpaši netraucēs. Šīs filiāles ir šādas:
13 22 14 10 13
14 11 14 11 12 11 28 — 1600±240 gadus pirms kopīgā senča (ir izceltas trīs mutācijas no uzrādītā haplotipa). Iesniegtais haplotips principā var būt jebkurā no šīm filiālēm; tā tikai 12 marķieri neļauj izslēgt nevienu no četrām filiālēm. Šo četru zaru kopējie senči dzīvoja, kā norādīts pa labi no senču haplotipiem. Apskatīsim, kurām apakškādēm pieder haplotipi, kas saistīti ar to, kas parādīts ar iepriekš minētajiem skaitļiem, un kuros pilnībā atkārtojas tā pirmie 12 marķieri. Šī materiāla beigās ir sniegti genoma dati par četriem no šiem indivīdiem, kuros ir atzīmēti SNP lejpus M253. Visticamāk, ID00020 būs tādi paši snips. Kopumā lielākajai daļai šo haplotipu ir SNP: I1-M253 > DF29 > CTS6364 > M227 un citā nozarē I1-M253 > DF29 > Z58 > Z138. ID00020 izgriezumiem ir jāpieder vai nu pirmajai, vai otrajai ķēdei. Tie visi tiks noteikti Maskavas DNS ģenealoģijas laboratorijā. Vēsturiskais ceļš uz iesniegto haplotipu ir aptuveni šāds. I haplogrupa izveidojās apmēram pirms 43 tūkstošiem gadu (no apvienotās haplogrupas IJ), apmēram pirms 28 tūkstošiem gadu tā izšķīrās haplogrupās I1 un I2 (lai gan “atšķīrās” šeit ir relatīvs termins, tie neatšķīrās, tie bija neatkarīgi notikumi) , no pirmās (tāpat kā no otrās) laika gaitā izveidojās daudzas apakšklādes. Haplogrupa I tika atrasta pārakmeņotās skeleta paliekās, kas datētas ar 7000 gadiem Zviedrijā un Centrāleiropā. Intervālā pirms 4600-4000 gadiem, kad Erbins apmetās Eiropā, gandrīz pilnībā izzuda haplogrupas R1b, haplogrupas I1 nesēji no Centrāleiropas, taču kāds izdzīvoja, un tā rezultātā mūsdienu I1 haplogrupas nesējiem ir salīdzinoši nesenais kopīgais sencis, kas radīja izdzīvojušos pēcnācējus aptuveni pirms 3700 gadiem, tas ir, 2. tūkstošgades pirms mūsu ēras pirmajā pusē. Tā kā haplogrupā I1 ir tikai viens kopīgs sencis, haplogrupas haplotipa koks ir tik simetrisks. Tas notiek ļoti reti. ID00020 ir šī kopīgā senča pēctecis. Pieteikums. Šeit mēs sniedzam detalizētus genoma datus par SNP sastāvu, kas atrodami ID00020 filiāles haplotipu nesējos. Tam vajadzētu būt gandrīz tādam pašam, izņemot, iespējams, termināļa izgriezumus. Bet varbūt tie ir vienādi, kā vairākos no iepriekš minētajiem gadījumiem. SNP no M223 un zemāk ir izcelti: Komplekts Nr. 38184 (Matthew Hamilton Butcher, Amerikas indiānis, bet saņēma Y-DNS no eiropiešu, 110. haplotips iepriekš minētajā filiālē): DF29+, M227+, CTS10058+, CTS10140+, CTS10338+, CTS10834+, CTS11036+, CTS11042+, CTS11441+, CTS11526+, CTS11552+, CTS11552+, CTS11552+, CTS1014+1, CTS1+1, 11775+1,3 CTS2193+, CTS2375+, CTS2514+, CTS2524+, CTS2644+, CTS2738+, CTS3517+, CTS3654+, CTS3843+, CTS4088+, CTS4130+ , CTS4295+, CTS4437+, CTS4848+, CTS4982+, CTS5167+, CTS5408+, CTS5513+, CTS565+, CTS5650+, CTS5705+, CTS571+, CTS5783+, CTS571+, CTS5783+, CTS571+, CTS5783+, CTS571+, CTS5783+, CTS5+9,5+8 93+, CTS6140+, CTS6221+, CTS623+, CTS6265+, CTS6395+, CTS641+, CTS6629+, CTS674+ , CTS6932+, CTS7267+, CTS7329+, CTS7831+, CTS7949+, CTS8333+, CTS8345+, CTS8394+, CTS8420+, CTS8716+, CTS88+, CTS8716+, CTS88+, CTS8876+, CTS88+, CTS8876+, 4,+8,+8,+8 09+, F3692+, F719+, L118+, L121+, L123+, L124+, L125+, L132+, L15+, L16+, L187+, L350+, L403+, L41+, L468+, L470+, L498+, L509+, L574+, L575+, L578+, L740+, L748+, L75+, L75+, L7+, L7+, 7+, 7+, 7+ 759+, L772+, L80+, L81+ , M168+, M170+ , M213+, M227+, M235+, M253+, M258+, M294+, M299+, M307+, M89+, M94+, P123+, P124+, P125+, P126+, P127+, P129+, P130+, P133+, P134+, P135+, P134+, P135+, P1+, P1+, P1+, P1+, P1+ , P141+, P143+, P145+, P146+ , P148+, P149+, P151+, P157+, P158+, P159+, P160+, P161+, P163+, P166+, P187+, P212+, P30+, P316+, LPP PF2608+, PF2611+, PF2615+, PF2747+, PF2748+, PF2749+, PF2770+ , PF3562+, PF3574+, PF3639+, PF3640+, PF3641+, PF3660+, PF3666+, PF3672+, PF3675+, PF3677+, PF3780+, PF3677+, PF3780+, PF3800, PF8+1, PF8+1, PF3800 3814+, PF3815+, PF3819+, PF3822+, PF3836+, PF3837+, PF6464+, PF6469+, PF6470+ , PF6477+ , PF6479+, PF6520+, V241+, V250+, YSC0000207+, YSC0000227+, YSC0000256+, YSC0000272+, YSC0000281+, YSC0000281+, YSC0000281+, M+4,+0, M+4,+0, M+4,+0 M72-, Z58-, Z63-, Z17694-, S2348-, S4442-, M3453- Komplekts Nr. 38737 (Lielmāja, Vācija, haplotips 744): CTS10058+, CTS10140+, CTS10338+, CTS10362+, CTS109+, CTS11036+, CTS11042+, CTS11358+, CTS16+1, CTS1+1,4+1,4+1,5+1,4 75+, CTS11726+, CTS11775+, CTS11783+, CTS11950+, CTS125+, CTS12632+, CTS1393+, CTS1996+, CTS2193+, CTS2375+, CTS2514+, CTS2524+, CTS2536+, CTS2644+, CTS2738+, CTS3331+, CTS3431+, CTS3517+, CTS3536+, CTS3517+, CTS3536+, CTS3517+, CTS3536+, CTS3+3, CTS3+8, CTS3+8, CTS3+8 68+, CTS39 96+, CTS4088+, CTS4130+, CTS4295+, CTS4364+, CTS4368+, CTS4443+, CTS4740+, CTS4848+, CTS4982+, CTS5167+, CTS5318+, CTS5408+, CTS5457+, CTS5513+, CTS5532+, CTS565+, CTS5650+, CTS5705+, CTS571+, CTS5705+, CTS571+, CTS5+9,83+, CTS5+9,5,8 +, CTS6135+, CTS6140+, CTS6221+, CTS6265+, CTS6383+, CTS6395+, CTS641+, CTS6629+, CTS674+ , CTS6800+, CTS6907+, CTS6932+, CTS7267+, CTS7329+, CTS7502+, CTS7831+, CTS7922+, CTS7933+, CTS7949+, CTS8243+, CTS8243+, CTS823+, CTS8+4, CTS833+4, CTS8333 8716+, CTS88+, CTS8876+, CTS8980+, CTS9240+, CTS9264+, CTS9288+, CTS9828+, F1046+, F1209+, F1302+, F1320+, F1329+, F1450+, F1460+, F1704+, F1714+, F1753+, F1767+, F2048+, F2075+, F2142+, F230+, F236+, F236+, F2352+, F2150+ +, F2408+, F2587+, F2688+, F2710+, F2794+, F2837+, F2985+ , F2993+, F3033+, F3111+, F3136+, F3335+, F3368+, F3402+, F3556+, F3692+, F4188+, F719+, F922+, L118+, L121+, L118+, L121+, L2,1+,1,1,1,5,1 , L16+, L187+, L350+, L403+ , L468+, L470+ , L498+, L509+, L574+, L575+, L578+, L740+, L748+, L75+, L750+, L751+, L755+, L756+, L758+, L759+, L772+, L80+, M3, M3, 1, 1, 1, 8 M253+ Z138+, Z139+, M170+, M258+, M307+, P19+, P30+, P38+, P109-, P259-, P37-, M161-, M72-, M26-, M21-, M223-, M227- Komplekts Nr. 152077 (Jens Pīters Nīlsens, Dānija, haplotips 745): CTS10058+, CTS10140+, CTS10338+, CTS10362+, CTS109+, CTS11036+, CTS11042+, CTS11042+, CTS11358+1, CTS11358+1, CTS11358+1,25, CTS1358+1, 25+1 C TS11575+, CTS11726+, CTS11775+, CTS11783+, CTS11950+, CTS125+, CTS12632+, CTS1393+ , CTS1996+, CTS2193+, CTS2375+, CTS2514+, CTS2524+, CTS2536+, CTS2644+, CTS2738+, CTS3331+, CTS3431+, CTS3517+, CTS3431+, CTS3517+, CTS36+, CTS3+, CTS3+, CTS3+, CTS36+,2 38 68+, CTS3996+, CTS4088+, CTS4130+, CTS4295+, CTS4364+, CTS4368+, CTS4443+, CTS4740+ , CTS4848+, CTS4982+, CTS5167+, CTS5318+, CTS5408+, CTS5457+, CTS5513+, CTS5532+, CTS565+, CTS5650+, CTS5705+, CTS571+, CTS5705+, CTS571+, CTS5705+, CTS571+, CTS5,9+9,9+9 93+, CTS6135+, CTS6140+, CTS6221+, CTS6265+, CTS6383+, CTS6395+, CTS641+, CTS6629+, CTS674+, CTS6800+, CTS6907+, CTS6932+, CTS7267+, CTS7329+, CTS7502+, CTS7831+, CTS7922+, CTS7933+, CTS7949+, CTS8243+, CTS7949+, CTS8243+, CTS7949+, CTS8243+, CTS4+8,833+, CTS4+8,833 0+, CTS8716+, CTS88+, CTS8876+, CTS8980+, CTS9240+, CTS9264+, CTS9288+, CTS9828+, F1046+ , F1209+, F1302+, F1320+, F1329+, F1450+, F1460+, F1704+, F1714+, F1753+, F1767+, F2048+, F2075+, F2142+, F2075+, F2142+, F23,+, F23,+, F23,+, F23,+ 02+, F2408+, F2587+, F2688+, F2710+, F2794+, F2837+, F2985+, F2993+, F3033+, F3111+, F3136+, F3335+, F3368+, F3402+, F3556+, F3692+, F4188+, F719+, F922+, F719+, F922+, L118+, L2, L2+, L2, L2, L2, L1+, L1 L157+, L16+, L187+, L350+, L403+, L468+ , L470+, L498+, L509+, L574+, L575+, L578+, L740+, L748+, L75+, L750+, L751+, L755+, L756+, L758+, L759+, L758+, L759+, L772+, L509+, L772+, M+1, 8 +, M253+, M294+, M307+, M42+, M450+, M89+, M94+, P123+, P124+, P126+, P127+, P130+, P135+, P136+, P138+, P14+, P141+, P4, P1+, P1+, P1+, P1+, P1+ P159+, P160+, P166+, P187+ , P30+, P305+, PAGES00026+, PAGES00081+, PAGES00123+, PF1016+, PF1029+, PF1031+, PF1040+, PF1046+, PF1061+, PF1061+, PF+1, PF+1, PF+1, PF+1, PF109 1269+, PF1276+, PF192+, PF210+, PF212+, PF223+, PF234+, PF258+, PF2591+ , PF2593+ , PF2599+, PF2608+, PF2611+, PF2615+, PF2624+, PF263+, PF2643+, PF272+, PF2745+, PF2747+, PF2745+, PF2747+, PF2748+, 7+PF28+7, PF2748+, 7+PF28 2+, PF316+, PF325+, PF342+, PF3515+, PF3517+, PF3518+, PF3534+, PF3560+, PF3561+ , PF3562+ , PF3574+, PF3578+, PF3586+, PF3588+, PF3590+, PF3594+, PF3596+, PF3600+, PF3604+, PF3605+, PF361+, PF+6,+1, PF361,+1,+3 PF3625+, PF 3639+, PF3640+, PF3641+, PF3642+, PF3654+, PF3660+, PF3666+, PF3672+ , PF3675+, PF3677+ , PF3686+, PF3694+, PF3780+, PF3800+, PF3804+, PF3806+, PF3807+, PF3809+, PF3811+, PF3811+, PF381, PF+8,+1, PF381,+8 PF3836+, PF3837+, PF 500+, PF667+, PF719+, PF725+, PF779+, PF796+, PF803+ , PF815+, PF821+, PF840+ , PF844+, PF892+, PF937+, PF951+, PF954+, PF970+, V168+, V186+, V189+, V205+, V221+, V205+, V221+, V20+, V2+0, V2+0, V2+0, V2+0, V2+0 56+, YSC0000256+, YSC00002 57+, YSC0000259+, YSC0000260+, YSC0000264+ , YSC0000265+, YSC0000267+, YSC0000272+, YSC0000280+, YSC0000281+, YSC0000298+, YSC0000299+, YSC0000300+, YSC000,0 Z138+, Z139+, M170+, M258+, M307+, P19+, P30+, P38+, P109-, P259-, P37-, M161-, M72-, M26-, M21-, M223-, M227- Komplekts N10060 (Robert Wade, Anglija, haplotype 57): M170+, M253+, M258+, M307+, P19+, P30+, P38+, P109-, P259-, P37-, M161-, M72-, M26-, M21-, M223-, M227- Haplogrupa I2a Diviem cilvēkiem bija šī haplogrupa, viena no apakšklases I2a1-L147.2, otra - I2a2-M223, ar attiecīgi 67 un 111 marķieru haplotipiem. KLIN ID00004 Tiek parādīts 67 marķieru haplotips, tiek ziņots par haplogrupu I2a. 13 24 17 11 14 15 11 13 12 13 11 30 – 17 8 10 11 11 25 15 20 0 12 14 15 15 – 10 10 20 21 15 12 19 17 35 35 11 10 – 11 8 15 15 7 12 10 8 11 9 12 22 22 16 10 12 12 12 7 10 30 21 13 14 10 13 11 11 12 9 Šī haplogrupa ir izplatīta visā Austrumeiropā, proti, Grieķijā, Serbijā, Bosnijā-Hercegovinā, Maķedonijā, Čehijā, Slovākijā, Polijā, Bulgārijā, Baltkrievijā, Krievijā, Ukrainā, Ungārijā, kā arī nedaudz Vācijā, Rumānijā un Itālijā un Lielākajai daļai pārbaudīto tika konstatēts I2a-CTS10228 SNP, kas pazīstams arī kā L147.2 (skat. ISOGG diagrammu zemāk; labajā pusē ir SNP veidošanās datējums saskaņā ar YFull uzņēmumu). Tas ir tipisks Austrumeiropas snips. Spriežot pēc haplotipa izskata, ir pilnīgi skaidrs, ka arī klientam tāds ir. Jautājums ir: vai mēs varam paredzēt dažus zemāka līmeņa SNP, pamatojoties uz haplotipa veidu?
Datu bāzē (FTDNA I2a Project) ir 245 haplotipi 67 marķieru formātā.No visiem šiem haplotipiem veidotā koka labajā pusē atrodas haplotips ID00004 (atzīmēts ar X), tas pieder pie saistītā lielā zara, kas aizņem lielāko daļu. labā, kompaktā zara koka. Kompakts nozīmē salīdzinoši jaunu zaru, salīdzinot ar irdenākiem zariem. Citiem vārdiem sakot, kompakts zars norāda uz tā salīdzinoši nesenu izcelsmi. Aprēķini liecina, ka šis zars ir aptuveni 2300 gadus vecs, tas ir, tas ir izveidojies pēdējās ēras beigās. Gandrīz visi šīs nozares haplotipi ir saistīti ar CTS10228/L147.2 snipi, taču tam ir arī lejupejoši snipi, piemēram, L147.2 >YP204/S17250 >Z16971 Kuri uz koka nav atdalīti 67 marķieru haplotipos. Precīzāk, iespējams, tie var atšķirties dažādās filiāles daļās, taču pārāk maz haplotipu nesēju projektā identificēja tik dziļus SNP. Tos noteiks Maskavas DNS ģenealoģijas laboratorija.
Redzam, ka uz 111 marķiera koka nav izteiktu zaru, ko varētu attiecināt uz konkrētiem cirpiem. Atzara pamata haplotips 67 marķieru formātā: 13 24 16 11 14 15 11 13 13 13 11 31 – 17 8 10 11 11 25 15 20 32 12 14 15 15 – 10 10 21 21 15 12 18 18 34 35 11 10 – 11 8 15 15 7 12 10 8 11 9 12 22 22 16 10 12 12 12 7 10 30 21 13 14 10 13 11 11 12 9 (Tika identificētas 8 mutācijas no iesniegtā haplotipa). Tas nozīmē, ka haplotips tiek noņemts no priekšteča par 8/0,12 = 67 → 72 konvencionālajām paaudzēm, tas ir, aptuveni 1800 gadus. Tas, protams, ir aptuveni, jo diviem haplotipiem precīzu aprēķinu nav - viena nejauša mutācija piedāvātajā haplotipā novirzīs aprēķināto attālumu par vairāk nekā 200 gadiem. Jebkurā gadījumā ir skaidrs, ka klienta senči ID00004 DNS līniju izsekojuši gandrīz tieši no šī atzara kopīgā priekšteča. Haplogrupas/apakšklādes I2a-L147.2 vēsture ar aptuveni 2300 gadu vecumu ir vairākkārt apspriesta vietnē Reformat. Haplogrupa I2a tika atrasta apbedījumos Centrāleiropā ar datējumu pirms 7000-5000 gadiem, pēc tam aptuveni pirms 4500 gadiem tā pazuda no Centrāleiropas un sadalījās divos zaros. Viens (precīzāk, vairāki zari, skatīt pirmo diagrammu iepriekš) pārcēlās (vai aizbēga) uz Britu salām, un šīs zaru kopas kopējie senči aizsākās apmēram pirms 4500-5000 gadiem. Cits apakšklādes I2a-L147.2 atzars izgāja cauri populācijas sašaurinājumam Eiropā, tika atjaunots aptuveni pirms 2300 gadiem Donavas-Karpatos un izplatījās visā Austrumeiropā mūsu ēras 1. tūkstošgades pirmajā pusē. Iespējams, vēsturnieki un arheologi to sajauca ar “slāvu izcelsmi”, un tas ir tas, kas par to tiek apspriests senajās hronikās, piemēram, stāstā par pagājušajiem gadiem. Ja PVL ir pareizi, tad I2a-L147.2 haplogrupas slāvi nākuši no Norikas, Alpu austrumos, uz ziemeļiem no Adrijas jūras ziemeļu daļas. Tauriskāni, Norikas iedzīvotāji, ir zināmi kopš 1. tūkstošgades pirms mūsu ēras otrās puses. Ja ID00004 būtu veicis BigY testu (vai citu Y hromosomas genoma testu), rezultāts būtu bijis tuvs (vai identisks) tālāk norādītajam (šeit ir izcelti I2a-CTS10228/L147.2 apakšklases augstākie SNP). Atlikušie iezīmētie SNP pieder vēl augstākiem haplotipiem, piemēram, piemēram, M42 - kombinētā haplogrupa VT; V9, V52 un V189 ir apvienota haplogrupa ST; M89 ir haplogrupa F, L15 un L16 ir apvienota haplogrupa IJK, L41 ir haplogrupa I utt. Kā redzat, vienīgā noderīgā lieta, ko šeit sniedza BigY tests (par USD 599), bija zemākā izgriezuma I2a-CTS10228 identificēšana. Šeit vairs nav pakārtotu izgriezumu, un iepriekš redzamais izgriezums Z17855 zemāk esošajā sarakstā ir negatīvs (ceturtais no beigām). Tālāk norādītais griezņu komplekts ir ņemts no atvērtas datu bāzes, tas pieder poļu prinča Svjatopolka-Četvertinska pēctecim, saskaņā ar leģendu - no poļu Rurikovičiem. Starp citu, kņazs Boriss Antonovičs Četvertinskis (1784-1865), Napoleona karu dalībnieks, Krievijas armijas pulkvedis, ir no tās pašas poļu ģimenes. Lielākā daļa, ja ne visi, no šiem izgriezumiem ir jāatrod ID00004. Tomēr tas viss ir tikai tad, ja dziļā apakšklāja izrādās tieši tāda pati kā Četvertinska. Bet interesanti ir tas, ka Četvertinskijā netika atrasti dziļi izgriezumi, kas būtu zemāki par L147.2; pats pēdējais termināls ir CTS10228, tas ir, L147.2. CTS4002+, CTS4039+, CTS4088+, CTS410+, CTS4437+, CTS4848+, CTS4982+, CTS5044+, CTS5375+, CTS5650+, CTS5727+, CTS5884+, CTS5727+, CTS5884+, CTS5727+, CTS5884+, CTS+6,5+9,5,9 +, CTS6265+, CTS674+, CTS6932+, CTS7175+, CTS7213+, CTS7218+, CTS7329+, CTS7831+, CTS8239+ , CTS8333+, CTS8345+, CTS8420+, CTS8486+, CTS88+, CTS8876+, CTS9240+, CTS9264+, CTS9349+, CTS10058+, CTS10228+, CTS10834+, CTS10936+, CTS11030+, CTS11441+, CTS11768+, CTS1293+, CTS176+, CTS1846+, CTS2193+, CTS2514+, CTS3296+, CTS2514+, FTS3296+, CTS+9,1,3+9,1,3 F3692+, F719+, L132+, L15+, L16+, L178+, L350+, L403+, L41+, L460+, L468+, L470+, L498+, L578+, M42+, M423+, M438+, M89+, M94+, P123+, P124+, P125+, P126+, P127+, P129+, P130+, P133+, P134+, P135+, P136+, P138+, P139+, P138+, P139+, P4+, P4+, P4+, P4+ , P146+, P148+, P149+, P151+ , P157+, P158+, P159+, P160+, P161+, P163+, P166+, P187+, P212+, P316+, P38+, PAGES00026+, PAGES00081+, PF2591+, 1+4, PF2591+, 1+4,PF259 7+, PF2748+, PF2749+, PF2770+, PF3562+, PF3574+, PF3626+ , PF3638+ , PF3639+, PF3640+, PF3641+, PF3644+, PF3651+, PF3652+, PF3657+, PF3658+, PF3660+, PF3664+, PF3660+, PF3664+, PF3666, PF3664+, PF3666, PF7+6,+3 PF3672+, PF 3675+, PF3677+, PF3780+, PF3800+, PF3801+, PF3802+, PF3804+, PF3809+ , PF3811+, PF3812+ , PF3814+, PF3815+, PF3819+, PF3820+, PF3822+, PF3836+, PF3837+, PF3849+, PF3876+, PF3849+, PF3876+, PF3966, PF3876+, PF3966,+4,4+4,+4 PF6470+, PF6477+, PF 6479+, PF6520+, L621+, L68+, L748+, L751+, L755+, L756+, L758+, L772+, M168+, M170+, M213+, M235+, M258+, M294+, M299+, V241+, V250+, YSC0000207+, YSC0000227+, YSC0000256+, YSC0000272+, M423+, Z1-7,1-9, Z7-9 -, Z2109-, M3453-, S2348-, S4442- , P61-, L584-, CTS3802-, DF41- Tā kā mēs runājam par Rurikovičiem, atcerēsimies, ka Krievijas muižniecības asamblejas prinču grupai ir haplogrupa N1c1, savukārt Četvertinskim (un klientam) ir haplogrupa I2a. Rurikam, ja tāda pastāvētu, varētu būt tikai viena haplogrupa. Tātad jautājums par daudziem Rurikovičiem joprojām paliek atklāts, ja viņiem visiem ir dokumentāls pamats apgalvot, ka viņi ir Rurikoviči. Cik man zināms, nevienam nav nekādu dokumentālu pierādījumu, taču ir labi zināmi ekspertu viedokļi par prinču ciltsrakstiem. Turklāt poļu ģenealogiem, iespējams, ir viens viedoklis, savukārt krievu ģenealogiem ir cits viedoklis. Viņi, iespējams, mēģina noskaidrot lietas savā starpā, taču DNS ģenealoģija sniedz viņiem jaunus datus, kas jāapsver. Iespējams, Maskavas DNS ģenealoģijas laboratorija varēs identificēt ID00004 pakārtotos SNP (saistībā ar CTS1028) un saprast, kuros reģionos tie galvenokārt tiek izteikti. Tad personīgo ģenealoģiju var vēl vairāk padziļināt. KLIN ID00019 Tiek parādīts 111 marķiera haplotips, tiek ziņots, ka haplogrupa I2b, apakšgrupa I2b1, izgriezums M223. 15 23 15 10 15 15 11 14 12 14 12 32 16 8 9 11 11 25 14 20 28 11 14 14 15 11 10 19 21 15 14 18 21 32 40 12 10 11 8 16 16 8 11 10 8 10 9 12 22 22 15 11 12 12 15 9 14 27 21 11 13 12 12 11 12 12 11 32 13 8 15 10 24 27 16 12 11 14 11 12 9 13 11 10 10 12 32 10 12 22 13 11 10 20 15 23 9 24 14 12 14 27 12 21 18 12 15 17 9 12 11 Iesniegtajos datos izmantota novecojusi 2010. gada nomenklatūra; kopš tā laika SNP M223 ir klasificēts kā apakšklase I2a2a (sk. diagrammu zemāk, saīsinātā veidā). Labajā kolonnā (gados, pirms gadiem) ir parādīts aptuvenais vecākā un dažu citu SNP (kas mūs šeit interesē) izcelsmes datējums, ko aprēķinājis uzņēmums YFull, izmantojot genoma datus.
Augšējā ceriņu krāsa norāda uz apakšklādi I2a-L147.2, kas ir visizplatītākā I haplogrupā Austrumeiropā - no Grieķijas līdz Baltijas valstīm, ieskaitot Krieviju, Ukrainu, Baltkrieviju, bijušās Dienvidslāvijas valstis; Sarkanā līnija izceļ uzrādītā haplotipa I2a-M223 apakšgrupu. Apakšklāde pati par sevi ir sena, tai ir 17 400 gadu, un, protams, tai ir zemākas apakšklādes, kuras mēs tuvāk identificēsim šajā interpretācijā. Apakšklādes, kurām varētu būt ID00019, ir izceltas dzeltenā krāsā, kā parādīts tālāk. Mēģināsim noteikt uzrādītā haplotipa vietu FTDNA projekta I haplogrupas I2 haplotipu 111 marķieru haplotipu vispārējā kokā. Lai to izdarītu, mēs ņemsim visus 63 projekta I2 haplogrupas 111 marķieru haplotipus, pievienosim tiem uzrādīto haplotipu ID00019 un izveidosim koku no iegūtajiem 64 haplotipiem, izmantojot profesionālu datorprogrammu (PHYLIP). Šī eksperimenta mērķis ir noskaidrot, ar kādiem haplotipiem un no kurām apakškādēm ieskauj prezentētais haplotips. Šo haplogrupas I2 111 marķieru haplotipu koks ir norādīts zemāk.
Uzrādītā haplotipa atrašanās vieta uz koka ir norādīta ar indeksu X. Šis ir atsevišķs, labi definēts zars, kurā ir septiņi pēc izskata līdzīgi, iespējams, saistīti haplotipi. Informācijai visa I2 koka pamata (senču) haplotips ir šāds 13 24 16 10 13 15 11 13 12 13 11 30 17 8 10 11 11 25 15 20 29 12 14 15 15 10 10 19 20 15 14 17 18 34 36 12 10 11 8 15 16 8 11 10 8 11 9 11 21 22 16 11 12 12 15 8 13 25 21 11 13 11 13 11 12 12 11 30 14 8 15 11 25 27 18 12 11 12 12 12 9 12 11 10 11 12 31 11 12 22 14 11 10 22 15 20 11 23 16 11 15 25 12 22 18 12 14 17 9 12 11 Piedāvātais haplotips no tā atšķiras ar 70 mutācijām. Tas ir līdzvērtīgs 70/0,198 = 354 → 537 parastajām paaudzēm vai aptuveni 13 425 gadiem starp tām (0,198 ir mutācijas ātruma konstante 111 marķiera haplotipiem, bultiņa ir atkārtotu mutāciju korekcija). Citiem vārdiem sakot, haplogrupas kopīgais sencis ir ārkārtīgi tālu laikā no ID00019. Visiem 64 haplotipiem ir 3906 mutācijas no bāzes haplotipa, tas atbilst laikam, kad dzīvoja šī haplotipa mūsdienu nesēju kopīgais sencis, un kas šajā gadījumā ir 3906/64/0,198 = 308 → 440 nosacītas paaudzes (katra 25 gadi, šim periodam tās tika kalibrētas mutāciju ātruma konstantes) jeb pirms 11000±1100 gadiem. Ir skaidrs, ka viņš dzīvoja ilgi pēc apakšklādes I2 veidošanās (apmēram pirms 27 500 gadiem, ko noteicis YFull snips), un tas norāda, ka haplogrupa I2 šķērsoja smagu populācijas sašaurinājumu apmēram pirms 11 000 gadiem, iespējams, saistīts ar ledus laikmetu Eiropā. . Klienta un haplogrupas I2 kopīgais sencis dzīvoja aptuveni (13425+11000)/2 = 12200 gadus atpakaļ, tas ir, pat pirms sašaurinājuma. Mēs redzam, ka uzrādītais haplotips atrodas tajā pašā atzarā ar haplotipiem ar numuru 39, 44-46, 51, 52. Šīs filiāles bāzes haplotips 15 23 15 10 15 15 11 13 12 14 12 32 15 8 10 11 11 25 14 20 28 11 14 14 15 11 10 19 21 14 14 17 19 33 39 12 10 11 8 15 16 8 11 10 8 10 9 12 21 22 15 11 12 12 14 9 13 27 20 11 13 12 12 11 12 12 11 31 13 8 15 11 24 27 16 12 11 13 11 13 9 12 11 10 11 12 32 10 12 22 13 11 10 20 15 22 9 23 14 12 14 28 12 21 18 12 15 17 9 12 11 Tas atšķiras ar 23 mutācijām (atzīmētas) no uzrādītā haplotipa, tas ir, haplotips šai nozarei tiek noņemts no senču haplotipa par 23/0,198 = 116 → 132 konvencionālajām paaudzēm (katra 25 gadi), tas ir, aptuveni 3300 gadus. Tajā pašā laikā visiem septiņiem zara haplotipiem ir 163 mutācijas no iepriekš parādītā pamata haplotipa, tas ir, šīs filiāles kopējais sencis dzīvoja 163/7/0,198 = 118 → 134 konvencionālās paaudzes, tas ir, 3350 ± 425 gadus. pirms. Mēs redzam, ka šīs divas vērtības sakrīt gandrīz ideāli (3300 un 3350 gadi, īpaši aprēķinu kļūdas robežās). Tas nozīmē, ka uzrādītais haplotips gandrīz ideāli iekļaujas šajā septiņu haplotipu atzarā un atbilst tam gan pēc haplotipa, gan attāluma no tā, gan pēc zara kopējā priekšteča aprēķinātā vecuma. Tātad mēs atradām haplotipa ID00019 filiāli. Apskatīsim, kurām apakšklasēm pieder saistītie (DNS ģenealoģiskajā nozīmē) haplotipi, kas numurēti ar 39, 44, 45, 46, 51 un 52. 39. haplotips (Anglija) ir vistuvāk uz koka uzrādītajam haplotipam. Viņam ir snip P95, kura vecums ir 3100 gadi (shēmā šī materiāla sākumā iezīmēts dzeltenā krāsā). Tas praktiski sakrīt ar apskatāmās nozares vecumu 3350±425 gadi. Tālāk seko 51. un 52. haplotipi (Skotija un Zviedrija), tie netiek ievadīti SNP, kas ir dziļāki par M223, piemēram, ID00019. Bet projekts tos definē kā SNP, kas nav augstāki par Z161, L801 un CTS6433. Pēdējiem trim šīs nozares haplotipiem, 44, 45 un 46 (Anglijā, Portugālē un Šveicē), ir attiecīgi SNP Z79, CTS6433 un CTS6433, kuru vecums ir 2400, 3800 un 3800 gadi. Tādējādi ID00019 atzars ir salīdzinoši jauns, un tajā esošie SNP atbilst vecumam, ņemot vērā, ka SNP vecums tiek noteikts ar noteiktu kļūdu. Rezumējot, jāsecina, ka ID00019 šķembām ir jāatrodas interpretācijas sākumā esošajā izgriezuma diagrammā dzeltenā krāsā norādīto izgriezumu diapazonā un, visticamāk, starp šķembām P95, CTS6433 un Z79. Maskavas DNS ģenealoģijas laboratorija plāno noteikt visus šos SNP. Vēsturiskais ceļš uz iesniegto haplotipu ir aptuveni šāds. I haplogrupa izveidojās apmēram pirms 43 tūkstošiem gadu (no konsolidētās haplogrupas IJ), apmēram pirms 28 tūkstošiem gadu tā izšķīrās haplogrupās I1 un I2, un no pēdējās (kā arī no pirmās) izveidojās daudzas zemākas apakšgrupas. Haplogrupas I2 turpmākā vēsture ir aprakstīta iepriekšējās interpretācijās; Rezultātā lielākā daļa I2a haplogrupas nesēju no Austrumeiropas pieder L147.2 apakšklādes lejupejošajiem atzariem, kuru kopējais sencis dzīvoja 1. tūkstošgades beigās pirms mūsu ēras, aptuveni pirms 2300 gadiem. ID00029 ir cita, paralēla atzara M223 pārstāvis, bet salīdzinoši nesenas apakšklādes, kuru kopējais sencis dzīvoja 2. tūkstošgades pirms mūsu ēras otrajā pusē, aptuveni pirms 3300 gadiem. Pieteikums. Šeit mēs sniedzam detalizētus genoma datus par SNP sastāvu, kas atrodami ID00029 filiāles haplotipu nesējos. Tam vajadzētu būt gandrīz tādam pašam, izņemot, iespējams, termināļa izgriezumus. Bet varbūt tie ir vienādi, kā vairākos no iepriekš minētajiem gadījumiem. SNP no M223 un zemāk ir izcelti. Komplekts Nr. 227059 (Anglija, 39. haplotips filiālē): CTS10057+, CTS10058+, CTS10100+, CTS10125+, CTS10362+, CTS109+, CTS11358+, CTS11441+, CTS11441+, CTS115, CTS11441+, CTS115, CTS1+1,25,25 +, CTS1977+, CTS1996+, CTS2193+, CTS2392+, CTS2514+, CTS2536+, CTS3296+, CTS3326+, CTS3331+, CTS3431+, CTS3517+, CTS3536+, CTS3654+, CTS3662+, CTS3868+, CTS3996+, CTS4039+, CTS4088+, CTS429+, CTS4088+, CTS429+, CTS429+, CTS4+4, CTS4+4,431+, CTS4+4 8+, CTS444 3+, CTS4740+, CTS4848+, CTS4982+, CTS5286+, CTS5318+, CTS5457+, CTS5532+, CTS5650+, CTS5727+, CTS5908+, CTS6135+, CTS6136+, CTS616+, CTS6265+, CTS6331+, CTS6383+, CTS674+, CTS6800+, CTS6907+, CTS6800+, CTS6907+, CTS6+7,32+, CTS6,7+3,7 +, CTS7682+, CTS7831+, CTS7865+, CTS7922+, CTS7933+, CTS7934+, CTS8243+, CTS8333+, CTS8345+ , CTS8420+, CTS8449+, CTS88+, CTS8876+, CTS8901+, CTS8980+, CTS9183+, CTS9240+, CTS9264+, CTS9266+, CTS9482+, CTS9782+, CTS9482+, FTS9782+, CTS9482+, FTS9782+, CTS9482+, FTS9782+, CTS9482+, FTS9782+, CTS+0,1+0,1+0,1 , F1320+, F1329+, F14 50+, F1460+, F1704+, F1714+, F1753+, F1767+ , F2048+, F2075+, F2142+, F2155+, F2302+, F2345+, F2366+, F2402+, F2587+, F2688+, F2710+, F2794+, F2837+, F2794+, F2837+, F23,+13, F23,+13, F2915+, F2915+ 35+, F3368+, F3402+, F3556+, F3692+, F4188+, F719+, F922+, L132+, L15+, L16+, L34+, L35+, L350+, L37+, L403+, L460+, L468+, L470+, L498+, L578+, L59+, L68+, L74+, L7+, L5+, L7+, L5+, L7+, L5+, L7+ , L800+, M139+, M168+, M17 0+, M223+, M235+, M294+, M42+, M89+, M94+, P123+, P124+, P126+, P127+, P130+, P135+, P136+, P138+, P14+, P141+, P145+, P1+, P1+, P1+, P1+, P1+, P1+, P1+, P1+ P160+, P166+, P187+, P214+ , P215+, P216+, P217+, P219+, P220+, P221+, P222+, P223+, P305+, P95+, PAGES00026+, PAGES00081+, PF1016+, PF1029+, PF1016+, PF1029+0,4+1, PF1029+1,+1 061+, PF1092+, PF1097+, PF110+, PF1203+, PF1269+, PF1276+ , PF192+ , PF210+, PF212+, PF223+, PF234+, PF258+, PF2591+, PF2593+, PF2599+, PF2608+, PF2611+, PF2615+, PF26, PF4+2, PF26, PF2+2, PF26 PF2745+, PF2747+, PF2748+, PF2749+, PF2770+, PF278+, PF292+, PF316+, PF325+ , PF342+ , PF3515+, PF3517+, PF3518+, PF3534+, PF3560+, PF3561+, PF3562+, PF3573+, PF3574+, PF3578+, PF3574+, PF3578+, PF3586+5, PF3586+5, PF3586+5,+5 3595+, PF 3596+, PF3600+, PF3604+, PF3605+, PF3607+, PF3611+, PF3612+, PF3616+ , PF3618+, PF3621+ , PF3623+, PF3625+, PF3626+, PF3634+, PF3639+, PF3640+, PF3641+, PF3642+, PF3644+, PF3642+, PF3644+, PF365, PF+6,+5, PF3+5, PF3+5, PF3658+, PF3660+, PF 3664+, PF3666+, PF3667+, PF3669+, PF3671+, PF3672+, PF3675+ , PF3677+, PF3686+, PF3694+ , PF3780+, PF3798+, PF3800+, PF3801+, PF3802+, PF3804+, PF3806+, PF3807+, PF3807+, PF+1, PF+1, PF+1, PF380,+1 PF3815+, PF3819+, PF3820+, PF 3822+, PF3836+, PF3837+, PF3849+, PF3876+, PF500+ , PF667+, PF719+, PF725+, PF779+, PF796+, PF803+, PF815+, PF821+, PF840+, PF844+, PF892+, PF937+, PF951+, PF937+, PF951+, V+1, PF951+, V91, V+1 +, V205+, V221+, V241+, V250+, V52+, V9+, YSC0000056+, YSC0000256+, YSC0000260+, YSC0000265+, YSC0000267+, YSC0000272+, YSC0000280+, YSC0000281+, YSC0000281+, YSC000020+0+0, YSC000020+0 Z161+, Z162+, Z163+, Z164+, Z165+, Z168+, Z170+, Z172+, Z174+, Z175+, Z176+, Z177+, Z178+, Z179+, Z181+, Z183+, Z181+, Z183+, Z181+, Z183+, Z186+, Z183+, Z186+7,18+7, 0057+, CTS10058+, CTS10100+, CTS10125+ , CTS10834+, CTS11441+, CTS1977+, CTS2193+, CTS2392+, CTS2514+, CTS3296+, CTS3326+, CTS3517+, CTS3654+, CTS4039+, CTS4+, CTS4,4,4,8+, CTS4,4,3 CTS4848+, CTS 4982+, CTS5286+, CTS5650+, CTS5727+, CTS5884+, CTS5908+, CTS6136+, CTS616+ , CTS623+, CTS6265+, CTS6331+, CTS674+, CTS6932+, CTS7329+, CTS7331+, CTS7682+, CTS7831+, CTS7865+, CTS7934+, CTS7865+, CTS7934+, CTS8333+, CTS4,+8, CTS8+8,+8 8+, CTS8876+, CTS890 1+, CTS9183+, CTS9240+, CTS9264+, CTS9266+, CTS9482+, CTS9782+ , F1209+, F3692+, F719+ , L132+, L15+, L16+, L181+, L34+, L35+, L350+, L36+, L37+, L403+, L41+, L460+, L468+, L9+, L4+, L8, L8, L8+, L4+ 48+, L751+, L755+, L756+ , L758+, L772+, L800+ , M168+, M170+, M213+, M223+, M235+, M258+, M294+, M299+, M42+, M438+, M89+, M94+, P123+, P1, P1, P2+, P1, P2+, P1 +, P130+, P133+, P134+ , P135+ , P136+, P138+, P139+ , P14+, P140+, P141+, P143+, P145+, P146+, P148+, P149+, P151+, P157+, P158+, P159+, P16+, P1+, P1+, P2+, P1+, P2+, P1+ 12+, P214+, P216+, P21 7+ , P218+, P219+, P220+, P221+ , P222+, P223+, P316+, P38+, P95+ LPP +, PF3641+, PF3644+, PF3651+, PF3652+, PF3657+, PF3658+, PF3660+, PF3664+, PF3666+, PF3667+, PF3669+, PF3671+, PF3672+, PF3675+, PF3677+, PF3780+, PF3800+, PF3801+, PF3802+, PF3804+, PF3802+, PF3804+, PF3809+, PF3804+, PF809+1,+1, PF3809+1,+1, PF3809 815+, PF 3819+, PF3820+, PF3822+, PF3836+, PF3837+, PF3849+, PF3876+, PF6464+, PF6469+, PF6470+ , PF6477+, PF6479+, PF6520+, V241+, V250+, YSC0000207+, YSC0000227+, YSC0000256+, YSC0000272+, YSC0000272+,28 Z161+, Z163+, Z164+, Z165+, Z168+, Z170+, Z174+, Z175+, Z176+, Z178+, Z179+, Z184+, Z186+, Z188+, P37-, S2348-, S4442 Komplekts Nr. 173448 (Anglija, 44. haplotips): Z77+, L800+, L801+, V218+, Z186+, Z78+, Z79+, L1198+, Z190+, Z185+, Z171+, Z166+, CTS10057+, CTS10058+, CTS10100+, CTS10125+, CTS10834+, CTS11441+, CTS2134+, CTS21441+, CTS2134+, CTS2193+, CTS2134, CTS2193+, CTS4, CTS23+, CTS4, CTS23+, C4+3 26+, CTS3517+, CTS3654+, CTS4039+, CTS4088+, CTS429+, CTS4348+, CTS4437+, CTS4848+ , CTS4982+ , CTS5286+, CTS5650+, CTS5727+, CTS5884+, CTS5908+, CTS6136+, CTS616+, CTS623+, CTS6265+, CTS6331+, CTS6433+, CTS674+, CTS6932+, CTS7329+, CTS7331+, CTS7682+, CTS7831+, CTS7865+, CTS7934+, CTS8333+, CTS8345+, CTS8420+, CTS8449+, CTS8420+, CTS8449+, CTS8449+, CTS8449+,+8,+8 3+, CTS9240+, CTS9264+, CTS9266+, CTS9482+, CTS9782+, F1209+, F3692+, F719+ , L104+ , L132+, L15+, L16+, L181+, L34+, L35+, L350+, L36+, L37+, L403+, L41+, L460+, L468+, L498+, L578+, L59+7, L578+, L59+7, L5, L+7, 4 +, L758+ , L772+, L800+ , M168+ , M170+, M213+, M223+, M235+, M258+, M294+, M299+, M42+, M438+, M89+, M94+, P108+, P123+, P124+, P125+, P126+, P127+, P129+, P127+, P129+, P13+, P3+, P1+, P3+, P3+, P3+, P1+ , P139+, P14+, P140+, P141+ , P143+, P145+, P146+, P148+, P149+, P151+, P157+, P158+, P159+, P160+, P161+, P163+, P166+, P187+, P212+, P1+, P2+, P2+, P2+, P2+, P2+, P2+ 20+, P221+, P222+, P223+ , P305+ , P316+, PAGES00026+, PAGES00081+, PF2591+, PF2608+, PF2611+, PF2615+, PF2624+, PF2747+, PF2748+, PF274+, PF2748+, PF274+7,PF27,+3 PF3626+, PF3639+, PF3640+, PF3641+, PF3644+, PF3651+, PF3652+, PF3657+, PF3658+, PF3660+ , PF3664+ , PF3666+, PF3667+, PF3669+, PF3671+, PF3672+, PF3675+, PF3677+, PF3780+, PF3800+, PF3801+, PF3801+, PF380, PF+1, 3+1 PF3812+, PF 3814+, PF3815+, PF3819+, PF3820+, PF3822+, PF3836+, PF3837+, PF3849+ , PF3876+, PF6464+ , PF6469+, PF6470+, PF6477+, PF6479+, PF6520+, PK1+, SRY10831+, V221+, V241+, V250+, V241+, V250+, YSC000,27+0,25+0 YSC0000272+, Z163+, Z164+, Z165+, Z168+, Z170+, Z174+, Z175+, Z178+, Z179+ , Z184+, Z185+, Z188+, Z190+, Z79+, L707+, M223+, M253-, M26-, M284-, M379-, P78-, P95-, S2348-, S4442- Komplekts Nr. 18187 (Portugāle, 45. haplotips): CTS674+, CTS6800+, CTS6907+, CTS6932+, CTS7329+, CTS7331+, CTS7502+, CTS7682+, CTS7831+, CTS7865+, CTS7831+, CTS7865+, CTS7831+, CTS7865+, CTS7,+7,3,+7,3 8243+, CTS8333+, CTS8345+, CTS8420+, CTS8449+, CTS88+, CTS8876+, CTS8901+, CTS8980+, CTS9183+, CTS9240+, CTS9264+, CTS9266+, CTS9482+, CTS9782+, CTS9828+, F1046+, F1209+, F1046+, F1209+, F1302+, F13, F13+, F13, F13+ 04+, F171 4+, F1753+, F1767+, F2048+, F2075+, F2142+, F2155+, F2302+, F2345+, F2366+, F2402+, F2587+, F2688+, F2710+, F2794+, F2837+, F2985+, F2993+, F3111+, F3136+, F3111+, F3136+, F336+, F336+, F336+, F3336+, F3336+ +, F4188+, F719+ , F922+, L132+, L15+, L16+, L34+, L35+, L350+, L37+, L403+, L41+, L460+, L468+, L470+, L498+, L578+, L59+, L68+, L748+, L751+, L755+, L756+, L755+, L756+, L758+, L756+, L758+, M1,8+,1,8,1,8 0+, M223+, M235+, M294+, M42+, M89+, M94+, P123+, P124+, P126+, P127+, P130+, P135+, P136+, P138+, P14+, P141+, P145+, P1+, P1+, P1+, P1+, P1+, P1+, P1+, P1+ P160+, P166+, P187+, P214+ , P215+, P216+, P217+, P219+, P220+, P221+, P222+, P223+, PF1016+, PF1029+, PF1031+, PF1040+, PF1046+, PF1040+, PF1046+, PF210+1,PF2+1, PF+1, PF1061 203+, PF1269+, PF1276+, PF192+, PF210+, PF212+, PF223+ , PF234+ , PF258+, PF2591+, PF2593+, PF2599+, PF2608+, PF2611+, PF2615+, PF2624+, PF263+, PF2643+, PF272+, PF2643+, PF272+, PF2643+, PF272+, 4,7, PF72+, 4,7, PF7 9+, PF277 0+, PF278+, PF292+, PF316+, PF325+, PF342+, PF3515+, PF3517+ , PF3518+, PF3534+, PF3560+, PF3561+, PF3562+, PF3573+, PF3574+, PF3578+, PF3586+, PF3588+, PF3590+, PF3590+, PF3594, PF+5,+5, PF359, PF3604+, PF3605+, PF 3607+, PF3611+, PF3612+, PF3616+, PF3618+, PF3621+, PF3623+ , PF3625+, PF3626+, PF3634+ , PF3639+, PF3640+, PF3641+, PF3642+, PF3644+, PF3651+, PF3652+, PF3654+, PF3652+, PF3654+, PF365, PF365, PF8+6,+6,+3 PF3666+, PF3667+, PF3669+, PF 3671+, PF3672+, PF3675+, PF3677+, PF3686+, PF3694+ , PF3780+, PF3798+, PF3800+, PF3801+ , PF3802+, PF3804+, PF3806+, PF3807+, PF3809+, PF3811+, PF3812+, PF3812+, PF381, PF+8, PF+1, 8+1, PF381, PF3822+, PF3836+, PF3837+, PF3849+, PF 3876+, PF500+, PF667+, PF719+, PF725+ , PF779+, PF796+, PF803+, PF815+, PF821+ , PF840+, PF844+, PF892+, PF937+, PF951+, PF954+, PF970+, V186+, PF970+, V186+, V20+0, V18+0, V25+0, V25+0 , YSC0000256+, YSC0000260+, YSC0000265+, YSC 0000267+, YSC0000272+, YSC0000280+ , YSC0000281+, YSC0000298+, YSC0000300+, Z161+, Z162+, Z163+, Z164+, Z165+, M170+, M223+, M258+, P38+, L801+, Z186+, Z76+, L1316+, CTS10057+, CTS10058+, CTS1058+, CTS101, CTS10+1, CTS101, CTS10+1 , CTS11358+, CTS11441+, CTS11575+, CTS11726+, CTS125+, CTS12632+, CTS1996+ , CTS2193+, CTS2392+, CTS2514+, CTS2536+, CTS3296+, CTS3326+, CTS3331+, CTS3431+, CTS3517+, CTS3536+, CTS3654+, CTS3654+, CTS366+, CTS3+9, CTS36+9, CTS366+9, CTS3662+, 4088+, CTS429+, CTS4314+, CTS4348+, CTS4364+, CTS4368+, CTS4443+, CTS4740+, CTS4848+ , CTS4982+ , CTS5286+, CTS5318+, CTS5457+, CTS5532+, CTS5650+, CTS5727+, CTS5908+, CTS6135+, CTS6136+, CTS616+, CTS6265+, CTS6265+, CTS6265+, CTS6265+, CTS6433+, F3692+, F719+, L104+, L132+, L15+, L16+, L181+, L34+, L35+, L350+, L36+, L37+, L403+, L41+, L460+, L468+, L460+, L468+, L5+7, L5+7, L8+6, L5+7, +, L756+, L758+, L772+ , L800+, M168+, M170+, M213+, M223+, M235+, M258+, M294+, M299+, M42+, M438+, M89+, M94+, P123+, P124+, P2+, P2+, P1+, P2+, P2+, P2+, P2+ P133+ , P134+, P135+, P136+, P138+, P139+, P14+, P140+, P141+, P145+, P146+, P148+, P149+, P151+, P157+, P158+, P159+, P160+, P161+, P163+, P1, P1+, P1, P1+, P1+, P1 +, P21 7+, P218+, P219+, P220+, P221+ , P222+, P223+, P316+, PAGES00026+, PAGES00081+, PF2591+, PF2608+, PF2611+, PF2615+, PF2747+, LPP2747+, LPF2748, PF2747+, PF2748, PF2747+, PF2748, 3833+, Z2198+, CTS8444+, CTS6497+, CTS10057+, CTS10058+, CTS10100+, CTS10125+, CTS10834+ , CTS11441+, CTS2134+ , CTS2193+, CTS2392+, CTS2514+, CTS3296+, CTS3326+, CTS3517+, CTS3654+, CTS4039+, CTS4088+, CTS408+, CTS43+, CTS43, CTS43,8 TS4982+, CTS5286+, CTS5 650+, CTS5727+, CTS5884+, CTS5908+, CTS6136+, CTS616+, CTS623+ , CTS6265+, CTS6331+, CTS674+, CTS6932+, CTS7329+, CTS7331+, CTS7682+, CTS7831+, CTS7934+, CTS8333+, CTS8345+, CTS8345+, CTS8420+, CTS8+8, CTS8420+8, CTS8420+8, CTS8420+8, CTS8420+ 01+, CTS9183+, CTS9240+, CTS92 64+, CTS9266+, CTS9482+, CTS9782+, PF3562+, PF3574+ , PF3626+, PF3639+, PF3640+, PF3641+, PF3644+, PF3651+, PF3652+, PF3657+, PF3658+, PF3660+, PF3664+, PF3660+, PF3664+, PF3666, PF3664+, PF3666, PF7+6,+6,+3 PF3672+, PF3675+, PF3677+, PF3780+, PF 3800+, PF3801+, PF3802+, PF3804+, PF3809+ , PF3811+, PF3812+, PF3814+, PF3815+, PF3819+, PF3820+, PF3822+, PF3836+, PF3837+, PF3849+, PF3876+, PF3849+, PF3876+, Z17+, Z1,7+, Z17+, Z1,7+, Z172+ 8+, Z179+, P19+, Z181+, Z183+, Z184+, Z186+, V22 1+ , V241+, V250+, YSC0000207+, YSC0000227+, YSC0000256+, YSC0000272+, Z163+, Z164+, Z165+, Z168+, Z170+, Z174+, Z170+, Z174+, Z17, PF8+, Z17+, Z17+ 469+, PF6470+, PF6477+, PF6479+, PF6520+, Z168+, Z170+, Z188+, Z7 7+, Z188+, F1209+, F3406-, M161-, Z190 -, Z63-, Z79-, L1317-, Z171-, S2348-, S4442-, Z185-, Z187-, L1201-, P30-,1 P3-, 8 -, PF4225-, DF5-, PF3292-, CTS661-, L1198-, Z190-, Z2059-, Z2084-, Z2074-, L1272-, ZS6-, ZS7-, ZS11-, ZS14-, ZS15-, ZS16-, ZS18-, ZS26-, L1290-, Z78-, Z79-, M284-, M379-, P78-, P95-, L380-, M26-, M307-, M72-, M227-, M253-, M21-, Z166- Komplekts Nr. B3411 (Šveice, 46. haplotips): L801+, CTS6433+, M223+, L1201-, Z190-, Z78-, CTS5332- Komplekts Nr. 23157 (Skotija, 51. haplotips): P19+, M170+, M258+, M223+, P19+, P38+, M284-, M379-, P78-, P95-, P30-, P37-, M227-, M253-, M26-, M307-, M72-, M21-, M161- Komplekts Nr. 174472 (Zviedrija, 52. haplotips): M284-, L126-, M379-, P78-, P95- Haplogrupa I2c KLIN ID00005 Tiek ziņots, ka I2c-L596 apakšklāde, dziļāka apakšklāde FTDNA, nav identificēta. 14 24 15 10 12 13 11 13 12 12 11 28 – 18 8 8 11 11 24 15 21 31 11 11 15 15 – 10 10 19 21 15 13 17 17 34 34 14 10 12 8 16 16 8 13 10 8 10 9 12 21 21 16 11 12 13 13 8 14 25 21 12 13 11 13 11 12 12 11 Tā kā apakšklāde netika noteikta dziļāk par I2c-L596, faktiski definēto apakšklādu ķēde I haplogrupā ir diezgan īsa: I-M170 > I2-M438 > I2c-L596. Identifikācija FTDNA tika veikta pareizi, par ko liecina uzrādītā haplotipa atrašanās vieta uz 67 marķieru haplotipu koka. Bet ir iespējamas šādas dilstošās apakšklases, kas vēl nav noteiktas:
Tagad par 9 haplotipu atzaru (trīs no Gruzijas [numurs 427, 451 un 452], divi no Baltkrievijas, pa vienam no Lietuvas, Austrijas un Turcijas). Šīs nozares pamata haplotips (ir iezīmētas mutācijas atšķirības no uzrādītā haplotipa) 14 24 15 10 12 13 11 13 11 13 11 29 – 18 8 8 11 11 24 15 20 32 11 14 15 16 – 11 10 19 21 15 13 17 19 34 36 12 10 12 8 16 16 8 12 10 8 10 9 12 21 21 16 11 12 13 14 8 13 25 20 12 13 11 13 11 12 12 11 Visiem 9 zara haplotipiem ir 123 mutācijas no šī pamata haplotipa, kas nozīmē, ka zara kopīgais sencis dzīvoja pirms 123/9/0.12 = 114 → 129 konvencionālajām paaudzēm jeb pirms 3225 ± 430 gadiem. Iesniegtais haplotips atšķiras no bāzes ar 17 mutācijām (vairākas mutācijas tiek skaitītas saskaņā ar īpašiem noteikumiem), un tas aprēķina kļūdas ietvaros dod aptuveni tādu pašu laiku no kopējā priekšteča. Tas liecina, ka ID00005 ir tiešs pēctecis no haplogrupas I2c atzara kopīgā priekšteča, kas dzīvoja ap 2. tūkstošgades beigām pirms mūsu ēras.
Lai saprastu, cik sarežģīta un traģiska bija L596 apakšklādes un tās apakšgrupu vēsture, mēs atzīmējam, ka pati L596 apakšklāde radās aptuveni pirms 21 300 gadiem (kā liecina tās SNP kāpnes Y hromosomā), taču tā gandrīz pilnībā izmira. Eiropas vēstures laikā, pēc tam atkal atdzimis no dažiem izdzīvojušajiem L596 nesējiem. Rezultātā apakšnozares ID00005 priekštecis dzīvoja aptuveni pirms 3225 gadiem, blakus esošā apakšnozare (septiņiem haplotipiem) diagrammā iepriekš - aptuveni pirms 2475 gadiem, augšējā apakšnozare (deviņiem haplotipiem) - pirms 2800 gadiem, un visi trīs zari. kopā - pirms 5950 ± 645 gadiem . Un genoma analīze rāda, ka apakšējā apakšlāde I2c1 tika izveidota pirms 15 500 gadiem; nākamā apakšklāde zemāk, I2c1a-L1251 – pirms 10 100 gadiem, par pārējo datu vēl nav. Mēs redzam, ka līdz šim ir atklāti trīs L596 zari, katrs datēts apmēram pirms 3200-2500 gadiem.
Tātad, apkoposim mūsu šodienas zināšanu stāvokli. I haplogrupa ir ļoti sena, izveidojusies pirms vairāk nekā 40 tūkstošiem gadu, kur joprojām nav zināma, bet, visticamāk, Eiropā. Apmēram pirms 30 tūkstošiem gadu tas šķīrās apakšklādēs I1 un I2, un pēdējais sadalījās apakšklādēs I2a, I2b un I2c-L596 aptuveni pirms 21 300 gadiem. Haplogrupas I un I2 tika atrastas senās fosilās kaulu paliekās Centrāleiropā un Zviedrijā, kas datētas pirms 7 tūkstošiem gadu. Nedaudz vēlāk dzīvoja trīs atzaru kopīgs sencis I2c-L596, kura pēcnācēji šobrīd dzīvo no Britu salām līdz Turcijai. Bet pirms aptuveni 4500 gadiem I haplogrupas nesēju, tāpat kā visu citu Eiropā, dzīve tika strauji izjaukta, un gandrīz visas haplogrupas (izņemot R1b) pazuda no Eiropas vai aizritēja atpakaļ uz Eiropas perifērajiem reģioniem, dažas devās uz Rietumāzija un Kaukāzs. Šīs haplogrupas kā atsevišķi pārstāvji iziet cauri ilgam un sāpīgam izdzīvošanas periodam, un izdzīvojušie sāk vairoties, veidojot kopīgus jaunu zaru senčus. Starp tiem bija apakšklādes I2c-L596 nesēji, kuri sāka “augt” pirms 3200–2500 gadiem, tas ir, 2. gadsimta beigās - 1. tūkstošgades vidū pirms mūsu ēras. Gandrīz tajā pašā laikā sāka atdzimt haplogrupa I2a, kuras kopējais sencis dzīvoja pirms 2300 gadiem Austrumeiropā, bet pirms aptuveni 5000 gadiem Britu salās. Nedaudz agrāk haplogrupa I1 sāka atdzimt ar izdzīvojušo kopējo priekšteci aptuveni pirms 3700 gadiem. Šajā sakarā I2c-L596 apakšklādei ir vissenākās saknes attiecībā pret mūsdienu šīs haplogrupas nesējiem - apmēram pirms 6000 gadiem. Priecē tas, ka L596 apakšējās apakšklases ir sadalītas atbilstošos haplotipu zaros (skatīt diagrammu iepriekš). Tas nozīmē, ka tad, kad Maskavas DNS ģenealoģijas laboratorijā tiks identificētas šīs apakšējās apakšklādes pietiekamā skaitā cilvēku (piemēram, piecdesmit), mēs varēsim veikt detalizētāku haplotipu piešķiršanu apakšklādiem un sīkāk izprast. apakšklādu dominējošā ģeogrāfija. Personīgais mtDNS interpretācijas tests Noteikti būs noderīgi paplašināt personīgo interpretāciju klāstu, iekļaujot sieviešu haplogrupas, un, lai tās pārbaudītu, var analizēt gan sieviešu, gan vīriešu mDNS. Fakts ir tāds, ka vīrietis mtDNS saņem asinīs no savas mātes, bet tālāk mtDNS nepārraida, jo pēc apaugļošanas (vai tās laikā) tiek iznīcināti spermas mitohondriji. Sperma satur apmēram duci mitohondriju, jo astes rotācijai ir nepieciešama enerģija, un tieši mitohondriji to nodrošina. Kad sperma ir sasniegusi savu mērķi, mitohondriji tiek izmesti. Vai atpazīstat vīriešus? Ļoti retos gadījumos mitohondriji joprojām izslīd cauri, taču tie ir sveši mitohondriji attiecībā pret topošās māmiņas ķermeni, un tas, kā likums, nebeidzas labi. Un tas bieži beidzas ar akūtām patoloģijām. Īsāk sakot, mtDNS tiek nodota no mātes meitām visā paaudžu ķēdē, desmitiem, simtiem tūkstošu gadu, miljoniem gadu utt. Un dēliem - tikai vienai paaudzei. Dēli Y hromosomu nodod saviem dēliem, mtDNS nenodod nevienam - ne dēliem, ne meitām, bet pārējās savas 22 hromosomas, rekombinantās, nodod visiem. Tāpēc es atkārtoju, ka mtDNS testus var veikt gan sieviešu, gan vīriešu DNS. Abos gadījumos rezultāts ir vienāds - mātes mtDNS un visas iepriekšējās mtDNS ķēdes identificēšana ar visām uzkrātajām mutācijām. Tas, ko tas dod, ir aprakstīts daudzās publikācijās un jo īpaši mums vispiemērotākajā formā - grāmatās “Internets. Pētnieka piezīmes" (2010), 368.-379.lpp., un nedaudz paplašinātā veidā - grāmatā "Slāvu izcelsme" (2013), 265.-284.lpp. Pirms sākam kampaņu par mtDNA masveida personisko interpretāciju, mēs piedāvājam izmēģinājuma versiju, ko izstrādājis viens no mūsu kolēģiem. Tā kā šī ir testa versija, tad interpretācijas autora vārds pagaidām tiek slēpts. Šajā posmā ir svarīgi, lai produkts tiktu uzskatīts par vismaz pieņemamu un vēl labāk par labu vai pat izcilu. Tātad tas ir domāts potenciālajiem klientiem, kā arī visiem lasītājiem. Tātad, kā testa iespēju, apsveriet tālāk norādīto.
Iesniegtie dati ļauj izsekot ģints vēsturei dziļumā no paleolīta (akmens laikmeta) tikai līdz pat vairākus tūkstošus gadu pirms mūsu ēras. Tas ir saistīts ar faktu, ka, pirmkārt, sieviešu mitohondriju DNS ir ievērojami īsāks ļoti mainīgu reģionu garums, un tāpēc tā ir daudz mazāk informatīva. Un, otrkārt, sniegtie dati nesatur pieejamo informāciju - piemēram, H1c apakšklāde, kurai, kā tiks parādīts zemāk, pieder uzrādītais mutācijas attēls, satur vairāk nekā 20 apakšgrupas. Tāpēc ģints vēsturi var rekonstruēt tikai ar lielāku tuvināšanas pakāpi, salīdzinot ar Y hromosomas datu interpretāciju. HVR1 un HVR2 atšķirību analīze ar RSRS parāda, ka iesniegtais haplotips pieder mitohondriju haplogrupai H1c. Iepriekš minēto iemeslu dēļ sniegtais paraugs sakrīt ar daudzu citu mtDNS datiem: familytreedna.com datubāzē ir haplotipu nesēji ar identiskām RSRS vērtībām HVR1 un HVR2, un lielākā daļa no tiem pieder ziemeļu un austrumu daļai. Eiropa, piemēram, mtDNA Anelai Sarockai (Lietuva). Haplotipa vēsture. Tiek uzskatīts, ka mātīšu haplogrupa H ir radusies Rietumāzijā pirms aptuveni 30 tūkstošiem gadu, Eiropā ieradusies aptuveni pirms 20-25 tūkstošiem gadu un izplatījusies uz kontinenta dienvidrietumiem Francijas-Kantabrijas reģionā, kas, visticamāk, atbilst Madlēnas kultūrai. . Pēdējā ledāja maksimuma laikā pirms 20-13 tūkstošiem gadu lielākā daļa paleolīta apmetņu Ziemeļeiropā un Centrāleiropā izmira, un tāpēc H haplogrupas pārstāvji lielākā mērā izdzīvoja tikai Spānijas ziemeļos (tāpēc šobrīd šī haplogrupa ar augsts, vairāk nekā 50%, sastopams starp baskiem, kuri veidojās šajā reģionā, asimilējot Ķīnas un Kaukāza citplanētiešus no vietējās autohtonās populācijas). Šī (fosilā) haplogrupa tika identificēta La Pasiega un La Chora (Madeleine) alās. Vēlākos apvāršņos jāatzīmē agrīna haplogrupas klātbūtne Ziemeļeiropā un Krievijas līdzenumā, sākot no mezoneolīta - Sertejā (Smoļenskas apgabals), kur Serteja VIII slānī tika atklāts H haplotipa nesējs. kombinācijā ar vīrišķo haplotipu R1a1 (IV tūkstošgadē pirms mūsu ēras).pmē.). Šo arheoloģisko kultūru var uzskatīt par Piltuves biķeru kultūras tālāko perifēriju. Tāpat līdzīgi haplotipi tika atrasti apbedījumos 5.-4. gadu tūkstotī pirms mūsu ēras. Doņeckas apgabalā un Južnija Oļenija Ostrova apbedījumā (kur tika identificēta arī vīriešu haplogrupa R1a1*). Apakšklāde H1c. T477C indikators ļauj mums noteikt, vai haplotips pieder H1c apakškladei. Tiek pieņemts, ka apakšklašu H1, H3, kā arī māsas haplogrupas V izplatība ir saistīta ar Eiropas iekšējo ekspansiju Francijas un Kantabrijas reģionā pēc pēdējā ledāja maksimuma apmēram pirms 13 tūkstošiem gadu. Haplogrupa H1 veido ievērojamu Rietumeiropas mitohondriju DNS daļu, un to lielā mērā dala arī Spānijas baski (28%), portugāļi (26%), Andalūzijas iedzīvotāji (24%) un Paciego etnogrāfiskā grupa Kantabrijā. (24%). Ziemeļāfrikā Lībijas tuaregu vidū ir neparasti augsts H1 haplogrupas īpatsvars (61%), kas arī norāda uz reģionu, kas robežojas ar Spāniju. Arī mtDNS haplogrupa H1 ir izplatīta starp citiem Ibērijas pussalas, Ziemeļāfrikas un Sardīnijas iedzīvotājiem. Turklāt tās pārvadātāji veido vairāk nekā 10% iedzīvotāju Francijā, Britu salās, Alpos, daudzos Austrumeiropas reģionos un vismaz 5% citviet Eiropā. Haplogrupas H1c vecums ir aptuveni 9400 gadi. Un šajā sakarā rodas jautājums par viņa pārvietošanās laiku un datumu uz Ziemeļeiropu un Austrumeiropu. Šo apakšlādi atklāja Skandināvijas piltuves biķeru kultūras lauksaimnieki 3500.–2500. gadā pirms mūsu ēras. (Gokhem2). Līdzīgs haplotips netika atrasts iepriekšējās Skandināvijas un Ziemeļeiropas kultūrās. Ņemot vērā šīs apakšlādes parādīšanos Eiropas ziemeļos un austrumos jau mezoneolītā, plašā joslā no Oleny salas līdz Ukrainas dienvidiem, tas ļauj izvirzīt jautājumu par migrāciju no dienvidiem. Tā kā Francijas-Spānijas reģionu var uzskatīt par vienu no šīs haplogrupas izplatības centriem iepriekšējā laikmetā, par loģisku hipotēzi par šī haplotipa parādīšanos var uzskatīt versiju par iespiešanos Ziemeļeiropā un Austrumeiropā pa jūru no Sirds keramikas kultūras nesēju Vidusjūras baseins (D. L. Gaskeviča hipotēze par Ziemeļpontikas impresso). Tas ļauj saistīt H1 nesēju parādīšanos ārpus Rietumeiropas ar Cardiac keramikas kompleksa migrācijām (vai citām kultūras un arheoloģiskajām saitēm), kuru pārstāvji varētu pārvietoties gar krastu ar ādas laivām. Gaskevičs pat piesauca megalītu kultūru un kardiālās keramikas veidotājus no Rietumeiropas un Vidusjūras, kuru vidē šī subklāde ir sastopama, izmantojot to, kas pastāvēja līdz 6. gadu tūkstotim pirms mūsu ēras. Bosfora šaurums uz Krimu un Ukrainas dienvidiem caur Bug-Dņestras kultūru.
Analīzes rezultātā varam konstatēt, ka uzrādītais haplotips atbilst Funnel Beaker kultūras pārstāvja apakšklādei, kas atrasta Gokhem2, ko daudzi pētnieki uzskata par indoeiropiešu. Mātes haplogrupa H tika atrasta senās vīriešu haplotipu R1a1 (Dienvidu Olenijs Ostrovs, Serteja) klātbūtnes apgabalā, kas ļauj uzskatīt šo haplogrupu, kas ir dzimtene Krievijas līdzenumā un sava veida vīriešu R1a1 “biedru”. Ilgāki savienojumi meklēja H1c saknes Vidusjūrā. Lai detalizēti izpētītu klienta ciltsrakstu, ieteicams veikt detalizētāku mašīnrakstīšanu, jo, atgādinām, haplogrupa H1c satur vairāk nekā 20 dažādus apakšklases. Šādu tipēšanu drīz veiks Maskavas DNS ģenealoģijas laboratorija. Haplogrupa I2 radās Dienvidaustrumeiropā pirms vairāk nekā 15 tūkstošiem gadu. Tā lielākā koncentrācija ir atrodama Bosnijā-Hercegovinā un Dienviddalmācijā (Horvātijā), un tā arī veido ievērojamu procentuālo daļu no visām dienvidslāvu populācijām.
Augstais I2a blīvums šajās zonās liek domāt, ka mūsdienu Horvātijas Adrijas reģions un Dināru Alpi bija dabisks patvērums paleopopulācijām, kas pēdējā ledus laikmetā pārnēsāja I2a. Haplogrupas I2a pārstāvji atradās Balkānu pussalas senāko kultūru pirmsākumi, piemēram: Chel Kladove, Lepenski Vir, Gura Baciului, Nea Nicomedia. Uz šī klana pamata acīmredzot attīstījās illīru un trāķu ciltis. Filiāle I2a bija viena no pirmajām, kas apdzīvoja Sardīniju. Haplogrupas filiāles:I2a1Haplogrupa I2a1(M26) pārstāv aptuveni 40% no visām vīriešu dzimtēm Sardīnijā. Turklāt tas ir zems vai mērens sastopams arī Basku zemē un Ibērijas pussalā, un tas bija 1,6% (1/64) Maķedonijā dzīvojošo albāņu izlasē un 1,2% (3/257) izlasē. Čehovs YSTR variācijas vecums M26 apakškladei ir 8,0 ± 4,0 tūkstoši gadu (Rootsi 2004). Tiek pieņemts, ka filiāle I2a izcēlās no I2 apmēram pirms 11 000 gadiem Balkānos, un filiāle I2a1 izcēlās no I2a apmēram pirms 8000 gadiem Sardīnijā vai kaut kur Eiropas piekrastē tās tuvumā. I2a2 Haplogrupu sadalījums I2a2 I2a2(snp M423) ir raksturīga Dienvidaustrumeiropas iedzīvotājiem. Vislielākais blīvums ir Dalmācijā (Horvātijā) un Bosnijā-Hercegovinā (>50%). Tiek pieņemts, ka šajā apgabalā Dināras augstienē šī atzara cēlusies no I2a apmēram pirms 7500 gadiem. Visizplatītākā haplogrupa I2a2 sastopams starp slāvu tautām (īpaši dienvidslāviem), kā arī starp rumāņiem, moldāviem, ungāriem, dienvidu lietuviešiem, albāņiem, grieķiem, Itālijas ziemeļaustrumu, Anatolijas rietumu un Ziemeļkaukāza iedzīvotājiem. Augstākā haplogrupas frekvence un daudzveidība I2a2 Rietumbalkānu iedzīvotāju vidū liecina, ka mūsdienu Horvātijas Adrijas reģions kalpoja par patvērumu šīs haplogrupas nesējiem pēdējā ledāja maksimuma laikā. Snp P37.2 atzars radās apmēram pirms 10,7 ± 4,8 tūkstošiem gadu (Rootsi 2004). Snp P37.2 atzara vecums ir 8,0 ± 4,0 tūkstoši gadu (Rootsi 2004), bet snp M423 tas ir 8,8 ± 3,6 tūkstoši gadu (Underhill 2007). Peričiks uzskata, ka šī atzara izplatība notika “ne agrāk par holocēna pāreju un ne vēlāk kā agrīnajā neolītā” (Pericic 2005). Otrais koncentrācijas centrs I2a2 kas atrodas mūsdienu Moldovā un aptuveni sakrīt ar tripiļu kultūras robežām, acīmredzot šī apakšhaplogrupa bija raksturīga šai kultūrai, un līdz ar indoeiropiešu ienākšanu tā pievienojās viņu genofondam, nepiedzīvojot tādu nomākšanu kā citi kultūras atzari. haplogrupa es Centrālajā un Rietumeiropā. I2b1Apakšhaplogrupu sadalījumi I2b1 sakrīt ar haplogrupas sadalījumu I1, izņemot Fennoskandiju, kas liecina par tās klātbūtni vismaz vienā no paleolīta patversmēm, kur atradās arī haplogrupa I1. Prombūtne I2b1 Fennoskandijā var būt saistīts ar to, ka haplogrupa I2b1 agrākās apdzīvotās vietas reģionā ietekmēja "dibinātāja efekts" un ģenētiskā novirze tās retuma dēļ, jo haplogrupa I2b1 veido mazāk nekā 10% no kopējās Y-hromosomu daudzveidības populācijā norādītajā apgabalā ārpus Lejassaksijas. Haplogrupu sadalījums I1 Un I2b1 salīdzinoši labi korelē ar ģermāņu valodu runātāju izplatības vēsturiskajām robežām. Haplogrupa I2b1 konstatēts vairāk nekā 4% iedzīvotāju tikai Vācijā, Nīderlandē, Beļģijā, Dānijā, Anglijā (izņemot Velsu un Kornvolu), Skotijā, Zviedrijas un Norvēģijas dienvidu galā, kā arī Normandijas, Menas, Anžu provincēs. un Perche Francijas ziemeļrietumos, Provansā Francijas dienvidaustrumos, Itālijas vēsturiskajos reģionos - Toskānā, Umbrijā un Latijā; kā arī Moldovā, Rjazaņas reģionā un Mordovijā. Šķiet diezgan iespējams, ka I1 un I2b1 klātbūtne mūsdienu Francijā, Anglijā un Itālijā, kā arī Austrumeiropā jau ir saistīta ar ķeltu un ģermāņu ekspansiju, un pirmsindoeiropas laikos šīs haplogrupas koncentrējās tikai ziemeļos. Eiropā. Viena no haplogrupas atzariem I2b1, proti I2b1a(snp M284), sastopams gandrīz tikai Lielbritānijas populācijā, kas var liecināt par tās ilgo pastāvēšanas vēsturi Britu salās. Interesanti, ka ar zemu haplogrupu biežumu I1 Un I2b ir sastopami vēsturiskajos Bitīnijas un Galatijas reģionos mūsdienu Turcijā, kur tos varēja atvest ķelti, kuri tur migrēja pēc Bitīnijas Nikomēda I uzaicinājuma. Haplogrupa I2b1 sastopams arī aptuveni 1% Sardīnijas iedzīvotāju. Tiek pieņemts, ka I2b izceļas no I2 Centrāleiropā, netālu no lēnām atkāpšanās ledāja malas apmēram pirms 13 tūkstošiem gadu, I2b1- no I2b vēl tālāk uz ziemeļiem, tagadējās Vācijas teritorijā, apmēram pirms 9 tūkstošiem gadu. Konkrēti Lielbritānijas filiāle I2b1a izcēlās no I2b1 apmēram pirms 3 tūkstošiem gadu. I2b2Haplogrupa I2b2 tika atklāta skeleta paliekās, kas atrastas Lihtenšteinas alā, bronzas laikmeta arheoloģiskajā vietā Vācijas centrālajā daļā, kur tika atrasti arī artefakti no Urn Fields kultūras. No 19 vīriešu atliekām alā haplogrupa I2b2 tika atrasta 13, R1b vienā un R1a divās. Jādomā, ka ala atradās toreizējās haplogrupas I2b izplatības epicentrā. Haplogrupa I2 ir visizplatītākā tēva izcelsme bijušajā Dienvidslāvijā, Rumānijā, Bulgārijā un Sardīnijā, kā arī galvenā ciltsraksts lielākajā daļā slāvu valstu. Tās maksimālās frekvences ir novērotas Bosnijā (55%, tai skaitā 71% Bosnijas horvātos), Sardīnijā (39,5%), Horvātijā (38%), Serbijā (33%), Melnkalnē (31%), Rumānijā (28%), Moldovā. (24%), Maķedonija (24%), Slovēnija (22%), Bulgārija (22%), Baltkrievija (18,5%), Ungārija (18%), Slovākija (17,5%), Ukraina (13,5%) un Albānija ( 13,5%). Ģermāņu valstīs tas ir sastopams 5 līdz 10% biežumā. I2 filoģenēzeJa esat iesācējs ģenētiskajā ģenealoģijā, lūdzu, skatiet mūsu Ievadu filoģenētikā, lai saprastu, kā lasīt filoģenētisko koku. I2 filoģenētiskais koks pēdējo 10 gadu laikā ir daudz attīstījies, un augstākā līmeņa apakšklādes tika pārdēvētas biežāk nekā jebkurai citai haplogrupai, izņemot R1b. Lai izvairītos no jebkādām neskaidrībām, ieteicams vēlreiz pārbaudīt definējošās mutācijas (SNP), lasot vecākus pētījumus vai atsaucoties uz konkrētu apakšklādi. Vairākas nelielas apakšklādes netika iekļautas vietas trūkuma dēļ.  Noklikšķiniet, lai palielinātu
Izcelsme un vēstureI haplogrupa ir vecākā lielākā haplogrupa Eiropā un, visticamāk, vienīgā, kas tur radusies (izņemot ļoti nelielas haplogrupas, piemēram, C1a2 un citu haplogrupu dziļās apakšgrupas). Haplogrupa IJ būtu ieradusies no Tuvajiem Austrumiem uz Eiropu pirms aptuveni 35 000 gadu, pēc tam drīz pēc tam attīstījusies par haplogrupu I. Tagad ir apstiprināts, ka pirmie Homo sapiens, kas kolonizēja Eiropu Aurignacian periodā (pirms 45 000 līdz 28 000 gadiem), piederēja haplogrupām CT, C1a, C1b, F un I. Tiek uzskatīts, ka I2 (M438/P215/S31) radās vēlā paleolīta laikā, aptuveni pēdējā ledāja maksimuma (LGM) laikā, kas ilga aptuveni no 26 500 līdz 19 000 gadiem. I2, iespējams, parādījās Rietumeiropā, lai gan precīzu tā izcelsmes reģionu nevar noteikt, jo paleolīta eiropieši bija nomadu mednieki-vācēji. Vecākais I2 paraugs, kas iegūts no arheoloģiskajiem skeletiem, ir 13 500 gadus vecs vīrietis no Grotte du Bichon (Šveicē), kas saistīts ar azīliešu kultūru (sk. Jones et al. (2015)). Viņa mātes izcelsme bija U5b1h. Uz 2016. gada oktobri ir pārbaudīti 15 mezolīta Eiropas Y-DNS paraugi. No tiem viens piederēja haplogrupai C1a2 (Spānijā), viena F (Vācijā), divas I* (Francijā) un sešas I2 (Luksemburgā un Zviedrijā), tostarp I2a1* (P37.2), I2a1a1a. (L672), I2a1b (M423) un I2c2 (PF3827). Viņu nēsātās mātes (mtDNS) līnijas bija U2e, U4, U5a1, U5a2 un U5b. Četri paraugi no Krievijas piederēja Y-haplogrupai J*, R1a1 (2x) un R1b1a. Tas liecina, ka jau pastāvēja zināma daudzveidība starp mezolīta Eiropas ciltīm, lai gan daudzas no šīm līnijām (C1a2, F, I*, J*) tagad ir ārkārtīgi reti sastopamas. Šķiet, ka haplogrupa I2a1 ir iznākusi no neolīta perioda kā liela uzvarētāja iemeslu dēļ, kas vēl nav skaidri. Anatolijas agrīnā neolīta genomu analīze, ko veica Mathisons et al. (2015) parāda, ka I2c (L596) jau bija sastopams Turcijas ziemeļrietumos pirms 8500 gadiem, un to asimilēja neolīta laika lauksaimnieki ceļā no auglīgā pusmēness uz Eiropu. No 2014. līdz 2016. gadam veiktie pētījumi atklāja, ka visas mezolīta Eiropas ciltsrakstus absorbēja neolīta laikmeta zemnieku vilnis. Tomēr I2a izcēlās kā visizplatītākais starp tiem, veidojot 16% no 69 pārbaudītajiem neolīta Y-DNS paraugiem. Tas nozīmē, ka tā ir otrā visizplatītākā tēva cilts pēc G2a, Anatolijas zemnieku sākotnējās ciltsraksta. Šķiet, ka I2a1 ir īpaši uzplaukusi Starčevo–Kőrss–Criş kultūrā (6000–4500 p.m.ē.) Dienvidaustrumeiropā, kur tā joprojām ir viena no galvenajām vīriešu dzimtām. I2a1 parādījās arī Printed-Cardium Pottery kultūrā (5000–1500 p.m.ē.) Vidusjūras rietumos. Mūsdienu sardīnieši un baski lielāko daļu sava genoma ir mantojuši no tiem neolīta laika zemniekiem no iespiestās kardijas keramikas kultūras. Lai gan līdz šim agrīnā vai vidējā neolīta paraugos ir atrasts tikai viens I2a2 (I2a2a-M223 no Spānijas), daudzi no tiem parādījās vara un bronzas laikmetā. Vara un bronzas laikmeta paraugi sastāv no viena I2a2, diviem I2a2a un viena I2a2a1 (CTS616) Spānijā, viena I2a2a1b1b2 (S12195) Krievijas dienvidos (Yamna kultūra), viena I2a2a1 (CTS9183) un viena I2a2a2aty2a (L2a2aty2a) Ungārijā. ), sešas I2a2b (L38) Vācijā (Unetice un Urnfield kultūras). Vara laikmeta I2a1 paraugi ietver I2a1a1 no Ziemeļitālijas (Remedello kultūra), I2a1 no Ungārijas (Vatjas kultūra). I2c2 tika atrasts arī Unetic kultūrā Vācijā. Haplogrupa I2a1 (P37.2)Haplogrupa I2a1 ir līdz šim lielākais I2 atzars un visciešāk saistīta ar neolīta kultūrām dienvidaustrumu, dienvidrietumu un ziemeļrietumu Eiropā.  Haplogrupa I2a1a1 (M26)I2a1a (M26, L158, L159.1/S169.1) bija zināms kā I1b2 līdz 2005. gadam, I1b1b 2006.–2007. gadā un I2a1 no 2008. līdz 2010. gadam. Tas ir sastopams visā Rietumeiropā un sasniedz maksimālo biežumu sardīniešu vidū (37,5%). un baski (5%), divi populācijas izolāti. M26 ģeogrāfiski ir ierobežots ar Britu salām, Zemajām zemēm, Franciju, Vācijas rietumiem, Šveici, Sardīniju, Sicīliju, Itālijas rietumu krastu, Ibērijas salu un Magribas Vidusjūras piekrasti. Vienīgie M26 negatīvie L160 mutācijai ir tikai Īrijā. I2a1a-M26, iespējams, bija viena no galvenajām Rietumeiropas megalītu kultūru dzimtajām līnijām neolīta un halkolīta periodos. I2a1a1a (L672) tika atrasts jau mezolīta Zviedrijā, kas nozīmē, ka I2a1a mezolīta periodā bija ļoti plaši izplatīts no Ibērijas līdz Skandināvijai. Vēlāk viņi būtu pārņēmuši lauksaimniecību, sajaucoties ar Tuvo Austrumu jaunpienācējiem. Haplogrupa I2a1b (M423)I2a1b (M423, L178) bija pazīstams kā I1b līdz 2007. gadam un I2a2 no 2008. līdz 2010. gadam. Galvenā apakšklāde, kas pārstāv vairāk nekā 90% no visām M423 līnijām, ir L621 un tās apakšklase L147.2. Pārējās apakšklases ir L41.2(ļoti reti) un L161.1(pārsvarā sastopams Vācijā un Britu salās). Haplogrupa I2a1b-L621Pārsvarā šī nozare ir sastopama slāvu valstīs. Tās maksimālās frekvences ir vērojamas dināru slāvu (slovēņu, horvātu, bosniešu, serbu, melnkalniešu un maķedoniešu) vidū, kā arī Bulgārijā, Rumānijā, Moldāvijā, Rietumukrainā un Baltkrievijā. Tas ir mazāk izplatīts arī Albānijā, Grieķijā, Ungārijā, Slovākijā, Polijā un Krievijas dienvidrietumos. I2-L621 (L147.2+) ir zināms arī kā I2a-Din(Dinarikam). Augstā I2a1b-L621 koncentrācija Rumānijas ziemeļaustrumos, Moldovā un Ukrainas centrālajā daļā atgādina par Kucuteni-Trypillian kultūras maksimālo izplatību (4800-3000 p.m.ē.). Neviens Y-DNS paraugs no šīs kultūras līdz šim nav pārbaudīts, taču, tā kā tas attīstījās kā Starčevo–Kőrss–Criş kultūras atzars, iespējams, ka I2a bija viena no tās galvenajām tēvu dzimtām, un varētu būt palielinājies dibinātāja efekts. tā biežums Kukuteni-tripīliešu kultūra bija visattīstītākā neolīta kultūra Eiropā pirms indoeiropiešu iebrukumiem bronzas laikmetā, un šķiet, ka tai bija intensīvi kontakti ar stepju kultūru pirms Jamnas paplašināšanās Balkānos un Centrāleiropā (skat. R1a vēsturi un R1b). No 3500. g. p.m.ē., Jamnas perioda sākumā Pontikas-Kaspijas stepē, kukuteniešu-tripiliešu tauta sāka paplašināties uz austrumiem, tagadējās Rietumukrainas stepē, atstājot savas pilsētas (tolaik lielākās pasaulē). un pieņem arvien nomadiskāku dzīvesveidu, piemēram, viņu Jamnas kaimiņi. Var viegli iedomāties, ka kukuteni-tripiliešus asimilēja Jamnas kaimiņi un viņi izplatījās kā minoritātes cilts līdzās haplogrupām R1a un R1b, virzoties uz Baltijas jūru līdz ar auklas izstrādājumu paplašināšanos. Alternatīvi, I2-L621 līnijas varēja dzīvot relatīvi izolēti no vispārējās protoindoeiropiešu sabiedrības kaut kur ap Ukrainu, Poliju vai Baltkrieviju, un, gadsimtiem un tūkstošiem gadu ejot, tās būtu sajaukušās ar pārsvarā R1a populācijām ap tām. Iegūtā amalgama būtu kļuvusi par protoslāvu priekštečiem. Mūsdienās I2a1 ir piecas līdz desmit reizes biežāk sastopams nekā G2a Dienvidaustrumeiropā, savukārt neolīta periodā G2a bija aptuveni četras reizes biežāk. Ar ko var izskaidrot šo pilnīgo apvērsumu? Šķiet, ka kādā vēstures posmā I2a1 ciltsraksti ir guvuši labumu no atrašanās uzvarētāju pusē. Neatkarīgi no neliela stimula (hipotētiska) pievienošanās Jamnas rietumu ekspansijai uz Eiropu, galvenais noteicošais notikums, kas ļāva I2a1b-L621 kļūt par galveno Austrumeiropas ciltsrakstu, iespējams, bija slāvu migrācija no 6. līdz 9. gadsimtam mūsu ēras. Lielākā daļa mūsdienu austrumeiropiešu kas pieder I2a1b, iekļaujas L147.2 (pazīstams arī kā CTS10228, CTS2180 vai Y3111) apakšklādē, kas, domājams, radās pirms 5600 gadiem (tieši pirms Jamnas perioda un Trypillian izplešanās stepē), bet kuras TMRCA ir tikai 2300 gados saskaņā ar Yfull. I2a1b-L621 indivīdu mazākums, kas negatīvi ietekmē L147.2, ir atrodami Polijas austrumos, Baltkrievijā un Ukrainas rietumos, kas liecina, ka šī cilts ir saglabājusies kopš halkolīta laikmeta. I2a1b-L147.2 apakšklāde, šķiet, ir ļoti strauji paplašinājušies, salīdzinot ar 1900 gadiem, kas atbilst slāvu etnoģenēzes laikam, ņemot vērā, ka paiet daži gadsimti, līdz vienam vīrietim var būt pietiekami daudz vīriešu kārtas pēcnācēju, lai tas sāktu ietekmēt populācijas mērogu. Šim I2-L147.2 priekštecim būtu tik liela ietekme uz plaukstošo agrīno slāvu populāciju, kas vēl bija neliela pirms 2300 gadiem, bet strauji attīstās. Pēc tam, kad Vācijas austrumos un Polijā dzīvojošās ģermāņu ciltis, piemēram, goti, vandaļi un burgundieši, iebruka Romas impērijā, tālāk uz austrumiem dzīvojošie slāvi aizpildīja vakuumu. Pēc Rietumromas impērijas sabrukuma 476. gadā slāvi pārcēlās uz Dināru Alpiem un Balkāniem. Līdz 9. gadsimtam slāvi ieņēma visas mūsdienu slāvu valodā runājošās teritorijas, izņemot Austrumbalkānus, ko kontrolēja turku valodā runājošie bulgāri. Mūsdienās ziemeļu slāvu valstīs ir no 9% (Polija, Čehija) līdz 21% (Ukraina) I2a-L621, savukārt dienvidu slāviem ir no 20% (Bulgārija) līdz 50% (Bosnija). Lielāks I2a-Din procentuālais daudzums dienvidos, iespējams, ir saistīts tikai ar citu dibinātāju efektu, kas saistīts ar faktu, ka dienvidslāvi cēlušies Ukrainas rietumos, kur I2a un R1a attiecība bija augstāka. Praktiski visi Dinaric I2a ietilpst L147.2 atzarā, un lielākā daļa ir S17250 atzarojums, kas cēlušies no kopīga patrilineāra senča, kurš dzīvoja tikai pirms 1800 gadiem. Haplogrupa I2a1b-L161.1Ģenētiskās ģenealoģijas aprindās plaši pazīstams kā I2-M423-Isles, L161.1 visbiežāk sastopams Rietumīrijā (5-10%) un Skotijas augstienēs (1-5%), bet zemās frekvencēs (> 1%) ir sastopams arī visā Centrāleiropā un Rietumeiropā, no Latvijas, Lietuvas un Baltkrievijas līdz Britu salām un no Skandināvijas līdz Spānijas ziemeļrietumiem. Tas ir atrasts arī Albānijā, Grieķijas ziemeļos, Bulgārijā un Rumānijā. Vecākie zināmie I2a1b-L161.1 indivīdi ir 8000 gadus vecs Loschbour vīrietis no mezolīta Luksemburgas un 7800 gadus vecs vīrietis no Motalas Zviedrijas dienvidos. I2a1b-L161.1, iespējams, bija izkaisīts pa lielāko daļu Eiropas vēlā ledāja un tiešā pēcleduslaika periodā un, visticamāk, integrēja neolīta sabiedrību tāpat kā visas pārējās mezolīta cilmes. I2a1b-L161.1, ļoti iespējams, bija viena no galvenajām neolīta līnijām Britu salās megalīta periodā, un tāpēc tā būtu bijusi starp cilvēkiem, kuri uzcēla Stounhendžu, Kūtu, Ņūgreidžu un citas lieliskas megalīta vietas. Tās zemais biežums mūsdienās un norobežojums Eiropas ziemeļrietumu un dienvidaustrumu nomalē neapšaubāmi ir daudzo indoeiropiešu migrācijas viļņu rezultāts pēdējo 5000 gadu laikā. Tagad zināms, ka protoķeltu haplogrupa R1b-L21 ieradās Īrijā ap 2000. gadu pirms mūsu ēras (sk. Cassidy et al. 2015), tikai dažus gadsimtus pēc R1b pirmās ierašanās Centrāleiropā. Pāreja bija strauja, R1b ciltīm tikai dažu gadsimtu laikā pārspējot Britu salu neolīta iedzīvotājus un virzot tās tālāk uz rietumiem un ziemeļiem, kur L161.1 saglabājas mūsdienās. Haplogrupas I2-L880, I2-L1286 un I2-L1294Šīs nelielās apakšklādes zemās frekvencēs ir sastopamas galvenokārt Ziemeļrietumu Eiropā. Tāpat kā I2a2 (skatīt zemāk), šīs līnijas, iespējams, ir Centrālās un Ziemeļrietumu Eiropas mednieku-vācēju pēcteči. L880 ir ļoti reta apakšklāde, kas identificēta Vācijā, Luksemburgā un Francijā. L1286 radās vēlā paleolīta periodā. Tomēr 99% mūsdienu nesēju pieder L233 apakšklasei, un tiem ir kopīgs patrilineārs sencis pirms mazāk nekā 2000 gadiem. L233 ir visizplatītākais Britu salās, bet reizēm sastopams arī Nīderlandē, Skandināvijā, Vācijā, Francijā un Polijā. L1294 ir arī ļoti reti sastopams un ir konstatēts tikai Vācijā, Francijā un Lielbritānijā. Haplogrupa I2a2 (P214)I2a2 (S33/M436/P214, P216/S30, P217/S23, P218/S32, L35/S150, L37/S153, L181) bija pazīstama kā I1c līdz 2005. gadam un I2b līdz 2010. gadam. Tas ir saistīts ar pirms ķeltu-ģermāņu Ziemeļrietumu Eiropas iedzīvotāji, piemēram, megalītu celtnieki(5000-1200 p.m.ē.). Tiek lēsts, ka tā vecums ir no 21 000 līdz 13 000 gadiem, kas atbilst epipaleolīta periodam. I2a2 ir sastopams lielākajā daļā Eiropas, un šķiet, ka tas bija izplatīts visā kontinentā pirms neolīta laika lauksaimnieku ierašanās. Līdz šim ir identificēti vairāki mezolīta I2a2 paraugi, galvenokārt Mathieson et al. (2017) Tas ietver personas no Vācijas dienvidiem (M223 no ap 7200. g. p.m.ē.), Dzelzs vārtiem starp Serbiju un Rumāniju (Z161 no ap 6200. g. p.m.ē.), Latvijas (CTS10057. no ap 5500. g. p.m.ē.) un Ukrainas dienvidaustrumiem (L699. no ap 5400. g. p.m.ē. un L701 no aptuveni 5200. g. p.m.ē.). Turklāt apakšklase I2c1 (L1251) nesen tika identificēts. Pārsvarā sastopams Vācijā un Anglijā un to perifērijā (Īrijā, Norvēģijā, Francijā, Itālijā, Polijā). Tiek uzskatīts, ka šī apakšklāde ir aptuveni 6000 gadus veca, kas to iedala vēlā neolīta periodā. To var saistīt ar medniekiem-vācējiem no Vācijas, kuri pārņēma lauksaimniecību pēc saskarsmes ar Tuvo Austrumu zemniekiem no lineārās keramikas kultūras. Slavenas personasI2a1b-L621 filiāle (slāvu)I2a2a-Z161 filiāle (vācu valodā)Endrjū Džonsons(1808-1875), Amerikas Savienoto Valstu 17. prezidents un 16. viceprezidents tika identificēts kā haplogrupas I2a2a (bijušais I2b1) dalībnieks, pamatojoties uz rezultātiem Presē Vispirms apskatīsim haplogrupu I2a1 (mutāciju P37.2). Slāviem (un gandrīz tikai slāviem!) raksturīgs tās atzars I2a1b (mutācijas L178/S32 un M423), precīzāk, apakšnozare I2a1b3 (mutācija L621/S392) un tā meita I2a1b3a (L147.2). Precīzāk vēl ir grūti pateikt, jo L621/S392 un L147.2 mutāciju klātbūtne ir pārbaudīta ļoti mazam indivīdu skaitam. Papildus slāviem starp tiem ir arī atsevišķi Vācijas, Itālijas un Lielbritānijas pārstāvji. Iespējams, ka starp 58 miljoniem nukleotīdu Y hromosomā tiks atklāta vēl vēlāka mutācija, kas raksturīga tikai slāvu apakšnozarei. Haplogrupas I2a1 slāvu apakšnozares izplatību labāk izsekot nevis pēc binārajiem marķieriem, bet gan pēc tai raksturīgajiem haplotipiem. Apakšnozarē I2a1b tie ir ļoti specifiski, ļaujot identificēt tās pārstāvjus ar augstu ticamību. FamilyTreeDNA (FTDNA) datubāze, kas nodrošina komerciālu testēšanu tiem, kas interesējas par DNS ģenealoģiju, satur visus šādas pārbaudes laikā identificētos haplotipus. Mūs interesējošā apakšnozare tur ir definēta kā “Dināra klasteris”. Tas sadalās divās daļās - dienvidu, faktiski dināriskajā, un ziemeļu, kas pārstāvēta visās slāvu populācijās (pareizāk to būtu saukt par "Karpatu"). Izmantojot šo datu bāzi, nav iespējams noteikt abu zaru biežumu dažādās populācijās, jo tajā uzrādītie paraugi tiek veidoti nejauši (vēlmes un iespēju pasūtīt DNS testu sakritība), ir ļoti dažādi pēc izmēra un nav reprezentatīvi. iedzīvotāju kopuma. Metodiski pareizi paraugi ir atrodami starptautiskajā Y-haplotipu (YHRD) atsauces datubāzē, kas izveidota un uzturēta kriminālistikas vajadzībām, kā arī līdzīgas nozīmes Baltkrievijas Republikas iedzīvotāju DNS marķieru atsauces datubāzē, kas bija izveidojis Baltkrievijas Republikas Tieslietu ministrijas Tiesu ekspertīžu un kriminālistikas centrs [Atsauces datubāze par Y-STR lokusu haplotipu sastopamības biežumu Baltkrievijā: http://dnkexpertiza.org/index1.html ]. Haplotipus dažiem paraugiem, kas nav iekļauti YHRD, var atrast zinātniskās publikācijās. Visus šos paraugus attēlo salīdzinoši neliels lokusu skaits, salīdzinot ar FTDNA: komerciālajos testos tiek izmantoti 37, 67 un pat 111 lokusu komplekti, bet tiesu ekspertīzēs labākajā gadījumā tiek izmantots 17 lokusu komplekts, kas pazīstams kā Y-Filer standarts. . Bet pat šis komplekts ļauj pārliecinoši atpazīt Dinaric klasterim raksturīgos haplotipus. Tiem ir raksturīgi salīdzinoši nelieli ģenētiskie attālumi no šīs klastera modālā haplotipa, kas Y-Filer formātā parādīts 1. tabulā. 1. tabula. Haplogrupas I2a1 dināru klastera modālais haplotips
Haplotipi, kas kopumā atšķiras no šī profila ne vairāk kā par 9-10 soļiem (atkārtojumu skaita mutācijas), vairumā gadījumu pieder pie mūs interesējošās I2a1 nozares. Iespējamā kļūda to noteikšanā nepārsniedz 1-2%, kas apstiprinās tajos (diemžēl dažos) gadījumos, kad paraugi tika vienlaikus pārbaudīti, izmantojot bināros marķierus. Dinaric klastera dienvidu atzars diezgan labi izceļas ar 67 un vēl jo vairāk 111 lokusiem, bet 17 lokusu Y-Filer komplektā vienīgā atšķirīgā iezīme ir samazinātais atkārtojumu skaits DYS-448 lokusā: 19 ( dažkārt pat 18), nevis 20. Protams, atšķirība starp diviem apakšklasteriem tikai vienu lokusu ir iespējama tikai kā pirmais tuvinājums, bet kopējā aina ir skaidra pat ar šādu aptuvenu novērtējumu. Haplotipu frekvences, kas, iespējams, pieder šim klasterim visiem publicētajiem Y-Filer formāta paraugiem, ir parādītas 2. 2. tabula. Dināru klastera haplotipu biežums
Tabulā redzams, ka haplogrupas I2a1 dināru klastera maksimums ir Balkānu-Karpatu reģionā, bet rietumu un austrumu slāvu vidū - Luka-Raykovets kultūras apgabalā. Gan Polijā, gan Krievijā tā biežums samazinās, bet Lietuvā un Vācijā tas ir tādā līmenī, ko var izskaidrot ar slāvu piejaukumu. Tas nozīmē, ka gan bastarniem, gan venēciešiem, kas atveda protoslāvu valodu no Lusatian-Pomožes apgabala, vēl trūka šīs kopas - tāpat kā senajiem baltiem pirms viņu intensīvo sakaru sākuma ar slāviem. Mūsdienu slāvu genofondā tas iekļuva ievērojamas autohtonu iedzīvotāju grupas asimilācijas rezultātā. No visām neslāvu tautām haplogrupa I2a1 ir augsta biežuma tikai starp rumāņiem, kas radušies agrīno slāvu (Prāgas un Ipotesti-čurēļu kultūras nesēju) sajaukšanas rezultātā ar vlahiem - trāķiešu pēctečiem. iedzīvotāju romanizēja Romas impērijas laikmetā. Turklāt biežums rumāņu vidū un slāvu etniskās grupas veidošanās apgabalā (Ļvovas apgabals, Baltkrievijas dienvidi) ir aptuveni vienāds. Līdz ar to vlahu vidū tas sākotnēji nebija zemāks par agrīnajiem slāviem. Dienvidu apakšklastera sadalījums ir indikatīvs. Tas parasti nav raksturīgs slāvu populācijām. Pat tie daži haplotipi ar DYS448=19, kas atrodas austrumu slāvu un poļu paraugos, ir diezgan tālu no modālajām vērtībām atlikušajiem lokusiem un, iespējams, atspoguļo neatkarīgas mutācijas. Tajā pašā laikā dienvidslāvu haplotipu profils bieži ir ļoti tuvu modālajam. Lielākā daļa viņu nesēju nepārprotami ir viena indivīda pēcteči, kuriem DYS448 lokusā bija 19 atkārtojumi. Šis indivīds dzīvoja vēlāk nekā visa Dināru klastera priekštecis, taču ne daudz - kopumā haplotipu izplatība dienvidu apakšklasterā ir diezgan liela, ģenētiskie attālumi 6-7 soļus no senču haplotipa tajā nav nekas neparasts. Šķiet, ka gan senču kopa ar 20 atkārtojumiem, gan meitas apakškopa ar 19 atkārtojumiem, līdz bronzas laikmeta beigām - dzelzs laikmeta sākumam, bija izplatīta galvenokārt uz dienvidiem no Karpatiem, turklāt meitas kopa radās dienvidu daļā. daļa no areāla un nebija laika izplatīties visā Karpatu baseinā pirms slāvu parādīšanās . Pēc Dacias evakuācijas 271. g. e. tas bija raksturīgi, acīmredzot, tikai bizantiešu subjektiem, kuri praktiski nešķērsoja Donavu. Tikmēr uz ziemeļiem no Karpatiem iekļuva vecāks un plašāks apakšklasteris ar DYS448=20, kur tas kļuva par daļu no topošajiem slāviem. Kad un kur slāvu senči iekļāva grupu, kurā bija liels “dinārisko” haplotipu īpatsvars? Tas nevarēja notikt agrāk par 2. gs. BC. - Zarubintsy kultūras veidošanās laiks. Bet tas nevarēja notikt vēlāk par 7.-8.gadsimta miju, kad atgriešanās migrācijas procesā uz ziemeļiem radās Luka-Raikoveca kultūra. Nevar citādi izskaidrot mūsdienu I2a1 frekvenci krievu un poļu vidū, kas veidojās tajās teritorijās, kur šīs haplogrupas sākotnēji nebija. Šeit ir divi iespējamie scenāriji. I2a1 nesēji varētu būt kādas vietējās kultūras pārstāvji, kas piedalījās Zarubincu kultūras veidošanā, vai kāda no vēlīnām Zarubincu grupām. Mēs varam runāt par milogradiem-neiriem, borisfenītiem-skitiem vai juhnoviešiem-melanhleniem. Otrajā scenārijā šī genofonda sastāvdaļa tika iegūta jau topošo slāvu apmešanās procesā, tiem asimilējot Čerņahovas kultūras paliekas Ukrainā vai romanizētos trāķus Karpatu baseinā un Donavas vidienē. Milogradas substrāta klātbūtne Zarubintsy kultūrā (īpaši tās Augšdņepras variantā) ir diezgan iespējama, lai gan dažādu pētnieku vērtējumi par tā lomu atšķiras. Vidusdņepras variantā skaidri pamanāms vēlo skitu substrāts. Nozīmīgs Juhnovska komponents piedalījās vēlo Zarubintsy grupu - Počepa, Kartamiševa un Grini-Abidnya tipa veidošanā, neskatoties uz to, ka pēdējiem bija izšķiroša loma Kijevas kultūras veidošanā. Bet šis scenārijs slikti izskaidro ziemeļu un dienvidu apakšklasteru haplotipu lielo līdzību. Tas, kas ir zināms par Juhnova kultūras izcelsmi, neļauj runāt par Balkānu-Karpatu reģiona kultūrām kopīgas sastāvdaļas klātbūtni tajā. Milogradas un mežstepju skitu kultūrās šāds komponents varētu būt izteikts dienvidrietumu impulss, kas fiksēts Černoles kultūras veidošanās laikā 1. tūkstošgades pirms mūsu ēras sākumā. e. Ar štancētās un rievotās keramikas prototrāķiešu radītājiem radniecīga populācija varēja “dināriskos” haplotipus atnest uz Ukrainas mežstepēm un pēc tam pēctecības kārtā nodot skitiem un milogradiešiem. Bet šo haplotipu nesējiem vairākus gadsimtus vajadzēja pastāvēt izolēti no galvenās Trāķijas pasaules, uzkrājot ievērojamu skaitu specifisku mutāciju. Tikmēr Baltkrievijas un Krievijas teritorijā nav īpašas I2a1 haplotipu specifikas. Otrajā, visticamākajā scenārijā ziemeļdināru haplotipu donori ir Karpatu pilskalnu kultūra, ko paplašināšanās procesā absorbējusi Prāgas kultūra (to identificē ar karpu cilti), un dako-romiešu atlikušās grupas. iedzīvotāju Karpatu baseinā. Kad slāvi sajaucās ar tiem Avar Khaganate ietvaros, ziemeļu subklādes biežums tika fiksēts aptuveni 20%. Tad viņi daļēji pārnesa savus haplotipus meža zonā, kur biežums samazinājās, sajaucoties ar vietējām populācijām. Tie, kas devās tālāk uz dienvidiem, Bizantijas impērijā, tur sastapa autohtonus, kuriem bija augsta dienvidu dināru haplotipu koncentrācija. Tieši šim substrātam horvāti, serbi un daļēji maķedonieši kopumā ir parādā augsto I2a1 frekvenci, kas ievērojami pārsniedz vispārējo slāvu līmeni. Slāvi, kas apmetās uz dienvidiem no Donavas, gandrīz nepiedalījās atgriešanās migrācijā, tāpēc meža zonā bija maz dienvidu dināru haplotipu. Vienīgie, kas šos haplotipus masveidā atveda uz Donavas ziemeļiem, bija austriešu slāvu senči, kā arī rumāņu priekšteči vlahi. Y-hromosomu haplogrupas I2a izcelsme, tās evolūcija un šīs haplogrupas nesēju populāciju migrācijas ceļi ir mana speciālā pētījuma priekšmets 5 gadus, sākot no brīža, kad sāku interesēties par aktīvu cilvēku populācijas izpēti. ģenētika. Iemesls manai interesei par šo haplogrupu ir pavisam vienkāršs - manas Y-hromosomas (Y-STR + Y-snips) ģenētiskā analīze parādīja, ka no filoģenētiskās klasifikācijas viedokļa tā pieder vienai no šīs hromosomas atzariem. haplogrupa, un šī banālā bioloģiskā fakta dēļ es cēlos no tiešās vīriešu līnijas no senajiem šīs haplogrupas pārstāvjiem. Mazāk svarīgs iemesls manai iesaistīšanai šīs haplogrupas izpētē ir nelaimīgais zināšanu trūkums gan par I2, gan par visu haplogrupu I. Pietiek ar to, ka pēdējie īpašie pētījumi par šo haplogrupu tika publicēti 2004., 2006., 2007. gadā - Rootsi , Sīri un citi.(2004), Rootsi, S. (2006), Peter Underhill et al (2007). Pārējie darbi, jo īpaši tie, kas attiecas uz populāciju Y-hromosomu daudzveidību Balkānos un bijušajā Dienvidslāvijā, galvenokārt atkārto iepriekšējos trijos darbos izteiktos secinājumus, neko nepievienojot (Marjanovičs, Damirs; et al., 2005, Peričić , Marijana un citi. 2005. gads; Rebala K., un citi.(2007). Tāpēc igauņu popģenētiķa pētnieka Rootsi darbs noteica vairākas pamathipotēzes par I haplogrupas izcelsmi, kuras vēlāk tika nostiprinātas zinātniskos un populārzinātniskos (Wikipedia) avotos. Diemžēl daudzas no šīm hipotēzēm izrādījās nepatiesas. Tālāk mēs iepazīstināsim ar dažiem noteikumiem, kas ietverti Vikipēdijas rakstā par I2, kas ir vispārpieņemti oficiālajā konservatīvajā populācijas ģenētikā, un, spriežot pēc raksta, daži no iepriekšminētajiem noteikumiem vēlāk tika pārdomāti un izstrādāti gandrīz ģenētikas amatieru aprindās.
Atstājot malā jautājumu par saikni starp haplogrupu un valodu, haplogrupu un arheoloģisko kultūru (šīs sakarības ir pelnījušas atsevišķu izpēti), redzēsim, ka Zviedrijas galvenais arguments par labu Balkānu senču dzimtenei I ir šīs haplogrupas ievērojamā klātbūtne Dinārā. Alpu/Balkānu reģions. Vienkāršākais un lētākais, no taupības viedokļa, skaidrojums šādiem skaitļiem ir pieņemt Balkānu populācijas genofonda daļas nepārtrauktību kopš pēdējā ledāja maksimuma Eiropā (pirms 20 tūkstošiem gadu), t.i., kopš Balkānu ledāju patvēruma laikiem. Kā zināms, Y-hromosomas haplogrupu ģeogrāfisko izcelsmi var noteikt pēc šāda empīriskā kritērija: vietā vai populācijā, kur radusies dotā haplogrupa, tās biežums un STR izkliede (vai STR mainīguma vecums) maksimums salīdzinājumā ar citām populācijām (Sengupta et al., 2006). Šī vienkāršā un šķietami loģiskā shēma, ja to aplūko sīkāk, izmantojot I haplogrupas piemēru, jauno datu gaismā neiztur kritiku. Tomēr ir vērts atcerēties, ka Rootsi rakstu iestatījums tika noteikts laikā, kad ģenētikas zinātniekiem bija tikai vispārēja izpratne par I haplogrupas struktūru. Turpmākie pētījumi, ko veica specializēti populācijas ģenētiķi, kopā ar īpašu amatieru pētījumu par I haplogrupu. komerciālie projekti FTDNA (Ģimenes koka DNS - Haplogroup I Y-DNA Project, FTDNA I1 yDNA Haplogroup, FTDNA I2*New ISOGG I2b, FTDNA I2a Y-Haplogroup, FTDNA I-M223 Y-Haplogroup, FTDNA I2a2b) un īpaši nozīmīgi-L centieni38a2b atsevišķu amatieru pētnieku pārstāvji tādā sauktā DNS ģenealoģijā (Kenets Nordtveds, Bernijs Kalens, Stīvs Trangsrūds, Viljams Hārtlijs, Īms Drosts, Ārons J. Hils, Pīters Hrečdakians, Hovans Simonians, Bobs Mejs, Vadims Verenihs, Bils Morovs, Lorenss Meika, Vladimirs Semargls u.c.) ļāva arvien labāk izprast haplogrupas zaru-apakšklādu filoģenētiskās attiecības un struktūras, kā arī nodrošināt hronoloģisku un ģeogrāfisku atsauci uz atsevišķiem haplogrupas zariem-apakšklādēm. Šis pētījums aprobežosies ar tikai īsu momentuzņēmumu par mūsu pašreizējo zināšanu stāvokli par haplogrupas I2a izcelsmi un “evolūciju”. Haplogrupas I2a izcelsme un migrācijas ceļi. Sakarā ar apskata fragmentāro raksturu, mēs neskarsim vienlīdz svarīgus jautājumus par to, kā un kad haplogrupa I parādījās Eiropā, kā arī jautājumus par I bazālā zara sadalīšanās laiku un vietu haplogrupās I1 un I2. Pašlaik mums nav pietiekami daudz pierādījumu par labu vienai no daudzajām un tikpat iespējamām scenārija versijām, un tāpēc mēs vienkārši apiesim šos jautājumus, cerot uz gaidāmo vidējā paleolīta Eiropas iedzīvotāju DNS izpēti. Tā vietā mēs sāksim aprakstu, prezentējot I2a haplogrupas filoģenētisko ('dzimtas koks') koku, atstājot malā citus šīs grupas zarus. Standarta vienkāršu I haplogrupas diagrammu vietā mēs piedāvājam koka kanonisko versiju Nordtvedt apstrādē (2013. gada versija). Tās galvenā priekšrocība ir haplotipu subklādu zaru un intraklādu kopu relatīvā topoloģiskā izvietojuma vizuāls attēlojums. Turklāt skala zem koka ir veidota, ņemot vērā zaru savstarpējās novirzes laiku (diverģences), ko aprēķina, izmantojot Kena Nordtvedta krusteniskās izkliedes metodi. Visbeidzot, filoģenētiskā koka grafisko attēlojumu papildina I haplogrupas nesēju kustības konjunktūras rekonstrukcija aizvēsturiskās Eiropas hronotopā. Haplogrupas I2a sadalījuma hronoloģija ir šāda (mūsu versija): Apmēram pirms 14 000 - 15 000 gadiem no plkst I2a L460/PF3647/S2381 ir parādījies zars I2a2-L35/PF3862/S150, L37/PF6900/S153, L181, M436/P214/PF3856/S33, P216/PF3855/S30, P217/PF3854/S23, P218/S3, kas gandrīz uzreiz sadalījās I2a2a-L34/PF3857/S151, L36/S152, L59, L368, L622, M223, P219/PF3859/S24, Paralēli tam no haplogrupas I2a parādījās apakšnozare I2a1-P37.2 (15 000–10 000 gadu pirms tagadnes). Jāatzīmē, ka paplašinātās analīzes laikā tika identificēta atsevišķa haplogrupa I2a1 -F (identifikācijas fragmenti L1294 L1297 L1298 nav iekļauti ISOGG kokā). Saskaņā ar Nordtvedt aprēķinātajiem starpklādu vecumiem šis klasteris parādījās vēlāk nekā I2a1a grupa, bet agrāk nekā I2a1b grupa (t.i., vidū starp abiem zariem). Saskaņā ar Nordtvedt, pēdējais kopīgais I2a1 sencis-F un I2a1a varēja dzīvot ne vēlāk kā 20 000 gadu pirms mūsu laika. Raksturīgi, ka I2a1*-F pārstāvji dzīvo galvenokārt mūsdienu rietumfrancijas teritorijā. Turklāt I2a1c pārstāvji dzīvo arī Rietumeiropā. No I2a1-P37.2no 20 000 līdz 15 000 pirms mūsu ēras kaste viņa laiks izcēlāsI2a1aL158/PF4073/S433, L159.1/S169.1, M26/PF4056((pēdējā kopīgā priekšteča mūžs — 8000–6000) unI2a1b-M423 L178/S328, M423(pēdējā kopīgā senča mūžs ir ne mazāks kā 14 000 gadu pirms tagadnes), no plkst I2a1b-M423 L178/S328 saskaņā ar interklādu aprakstu senču līnijas tika izdalītas apmēram pirms 13 500 gadiemI2a1b3 un I2a1b2 — (Dinaric+Disles) un salas. Interesanti, ka mēs lēšam, ka pēdējā kopīgā senča mūža ilgums salām ir aptuveni 6600 gadi pirms šī laika ir aptuveni 8000 gadu.
Vairāk par vēlīnās intraklādes dalīšanās (attieši 8000-6000 gadu pirms mūsdienām) labi korelē ar globālo atdzišanu apm. 6200. g. pirms mūsu ēras, kas izraisīja neolīta demogrāfisko krīzi, ko atzīmēja eksperti, kad iedzīvotāju skaits Eiropā nepārprotami samazinājās.Ziemeļeiropā tajā pašā laikā manā laikā (aptuveni 6500. - 6200. g. pirms mūsu ēras) notika notikums, kas ģeogrāfiski atdalīja (filoģenētiski atdalījās agrāk) populācijas I2a1b2-Isles un I2a1b3 , kā arī daļa no I2a2-M284 – Isles apakšklādes no atlikušajām I2a2 apakšklādēm.Mēs runājam par Dogerlendas plūdiem. Definēts arpierādījumi, tostarp mūsdienu jūras gultnes kontūras, liecina, ka pēdējā apledojuma laikā ūdensšķirtne starp Ziemeļjūru un Lamanšu virzījās no Austrumanglijas dienvidaustrumiem līdz Hoek van Holland, nevis caur Pas de Kalē, un ka Temzas upes, Mss, Šelds un Reina, apvienoti, plūda gar Lamanšu platas upes veidā, kas galu galā ieplūda Atlantijas okeānā.[Ap 8000.g.pmē. e. sauszemes ziemeļu krasts, Doggerland, bija piekraste ar lagūnām, purviem, plūdmaiņu krastiem un pludmalēm. Iespējams, mezolīta laikmetā šis apgabals bija bagātākais Eiropā medību, mājputnu un zvejniecības ziņā. Tiek pieņemts, ka pakāpeniska jūras līmeņa celšanās ledāja kušanas rezultātā (pēdējā apledojuma beigas) izraisīja Dogerlendas applūšanu pie Ziemeļjūras, kas ap 6500. gadu pakāpeniski atdalīja aizvēsturisko Lielbritāniju no Eiropas kontinenta. BC. e. Topošās smilšu sēkļa vietā, kas pašlaik pazīstama kā Dogger Bank, saskaņā ar šo hipotēzi sala turpināja pastāvēt vismaz līdz 5000. gadam pirms mūsu ēras. e. Alternatīva nesen izvirzītā hipotēze ir tāda, ka lielāko daļu zemes appludināja cunami aptuveni pirms 8200 gadiem (6200. gadus pirms mūsu ēras), ko izraisīja viens viens no lielākajiem vēsturē zemes nogruvumi - zemūdens augsnes laukuma novirzīšanās no krasta Norvēģija pazīstama kā Sturegga . Saskaņā ar šo teoriju zemes nogruvuma izraisītajam cunami bija katastrofāla nozīme apgabala mezolīta iedzīvotājiem. Šķiet, ka cunami rezultātā Lielbritānija ir neatgriezeniski atdalīta no kontinenta un no kultūras viedokļa Mezolīts gāja savu ceļu uz to. Viena no blakusparādībāmnotika zemes nogruvums globālā atdzišana 6200 BC e. aukstā ūdens pieplūduma dēļ no izkusušajiem ledājiem. Kas bija aizvēsturiskie I2a haplogrupas pārstāvji? es Kur parādījās haplogrupas I2a senči?? Kā redzams no iepriekš minētā Vikipēdijas citāta, dažās ģenētiskajās aprindās ir nostiprinājies, mūsuprāt, nepareizais viedoklis, ka I2a ir saistīts ar Balkānu neolīta kultūru loku, tostarp ar trimpiliešiem pietuvinātām kultūrām. Šī iespēja ir iespējama, taču galīgā versija jānosaka, analizējot Y-hromosomas, kas ir vecākas par šo kultūru pārstāvju mirstīgo atlieku DNS. Bet, ja I2 bija sastopams starp trypilliešiem, tos diez vai var uzskatīt par pamatiedzīvotāju elementu (kuru kodols bija imigranti no Tuvajiem Austrumiem), visticamāk, viņi pārstāvēja vietējā mezolīta elementa paliekas, kas iekļautas šajā kultūrā. Ja runājam par seno I2a2 pirmatnējo kultūru, tad tie visticamāk bija svīderieši un viņu epigoni. 2011. gadā man bija iespēja nedaudz apspriesties ar Polijas FTDNA projekta administratoru (Leriju Majku) par haplogrupas I2a1b-M423 (un I2a kopumā) izcelsmi. Pirms tam Maiku salīdzinoši maz interesēja šīs haplogrupas izcelsme (lai gan, tāpat kā es pats, viņš tai pieder). Viņš ierosināja ārkārtīgi interesantu versiju par haplogrupas I2a1 izcelsmes vietu. Pēc viņa domām, visu I2a1 (kā I2a1*, I2a1a, I2a1b, I2a1c) kopīgais sencis ar lielu varbūtības pakāpi varēja dzīvot Alpu reģionā. Viņš arī piekrita manam piedāvātajam maršrutam I2a1 klana pārejai no Alpiem uz Karpatiem. Tiesa, viņš manam randiņam nepiekrita. Uzskatu, ka I2a1 senču populācija sāka savu kustību no Alpiem uz Karpatiem (no Rietumeiropas uz ziemeļiem pēc ledāja atkāpšanās), tūlīt pēc LGM beigām (t.i., mezolītā kaut kur pēc 10 000. g. p.m.ē.), Maiks uzskatīja. ka migrācijas iniciatori varēja būt ķelti (tas ir, šī migrācija notika jau bronzas laikmetā). Pēdējais variants, lai arī interesants, neizskaidro salas apakšklādes I2a1b2 un dināru apakšklādes I2a1b3 atdalīšanas laiku un vietu, kas, pēc molekulārās datēšanas, nevarēja notikt vēlāk kā pirms 10-8 tūkstošiem gadu. Lai pārbaudītu Maika versiju, nolēmu izmantot Meskita algoritmu, kas ļauj novērtēt ticamāko (no taupības viedokļa) apakšklādes “pamata” vietu, pamatojoties uz pašreizējo ģeogrāfisko izplatību (ģeogrāfisko). izplatība) no divām blakus esošajām apakšklādēm (šajā gadījumā mēs nosakām parādīšanās vietu I2a1b1, pamatojoties uz haplotipa taksonu ģeogrāfisko koordinātu un I2a1b koka topoloģijas salīdzinājumu, kuru sakņoja ārpusgrupas sakne I2a1a). Neskatoties uz to, ka šobrīd vislielākais I2a1a apakšklādes blīvums ir Sardīnijā, saskaņā ar Nordtvedta pamatoto un empīriski apstiprināto viedokli vecākās I2a1a kopas atrodas ziemeļu reģionā. Ibērijas pussala, Alpi un daļa Francijas dienvidu. Šim nolūkam es uzzināju, kā Meskītā filoģenētiskos kokus pielietot tieši Ziemeļeiropas ģeogrāfiskajai projekcijai, 2D-3D projekcijā. Piemēram, 2 kartes, kuras es izveidoju programmā Mesquite Cartographer, pamatojoties uz filoģenētiskā koka struktūru ar 687 haplotipiem I2a1b-M423, kas tika identificēti, atrodot 20 neatkarīgas atbilstības starp 20 labākajiem (parsimonic) kokiem TNT. Sarkanā krāsa norāda uz rekonstruēto izcelsmes vietu – šajā gadījumā Alpu reģionu.
Tādējādi trajektorija, ko ieguvām I2a1b nesējiem, aptuveni atbilst mūsu piedāvātajam maršrutam. Grūti precīzi pateikt, kad notika migrācija, jo starp I2a1b un I2a1b3 parādīšanos pagāja vairāki tūkstoši gadu. Vēl viena lieta ir svarīga, mēs parādījām, ka kustība sākās nevis no Balkānu teritorijas, bet gan no Francijas vai Šveices Alpiem uz ziemeļaustrumiem. Vai šo versiju atbalsta lietiskie pierādījumi - senais DNS utt.? Burtiski līdz pavisam nesen mūsu ierosinātā hipotēze palika tikai statistiska interpolācija, kurai nepieciešams papildu apstiprinājums. Pirms pāris nedēļām beidzot tika publicēts ‘profesionālu’ ģenētiķu darbs, kas parāda, kā lielākā daļa līdz šim pētīto seno paraugu DNS korelē ar mūsdienu cilvēku populāciju DNS ( Senie cilvēka genomi liecina par trim senču populācijām mūsdienu eiropiešiem). Mans pieņēmums par I2a1b migrācijas sākumpunktu un sākumu, ko esmu paudis pēdējos gados, guva materiālu apstiprinājumu darbā. Pirmkārt, vienparentālajās līnijās Zviedrijas un Luksemburgas mezolīta mednieku-vācēju senais DNS gēnu fonds izrādījās mtDNS haplogrupas U un Y-hromosomu haplogrupas I pārstāvji, tāpēc atkal runa ir par R1a/R1b neesamību agrīnā stadijā. Mezolīta eiropieši. Otrkārt, šie pētījumi liecina juksemburgiešu Mezolīta mednieks-vācējs (kura mirstīgās atliekas tika atrastas Hefingenas-Ločburgas mezolīta vietā Luksemburgā aptuveni 8000 gadus pirms mūsdienām) piederēja haplogrupai. I2a1b* M423 + L178+. Saskaņā ar tabulā sniegtajiem datiem par Y-hromosomu polimorfismiem šīs aizvēsturiskās Luksemburgas haplogrupa ir senču grupa gan attiecībā pret I2a1b2, gan I2a1b3. I2a1b L178 G A 15574052 12+ Treškārt, šie pētījumi arī parāda, ka daļa Motalas (Zviedrija, Estgaterlande, aptuveni 8000 gadus pirms mūsdienām) mezolīta populācijas piederēja arī I2a1b*(xI2a1b1, I2a1b3) |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||






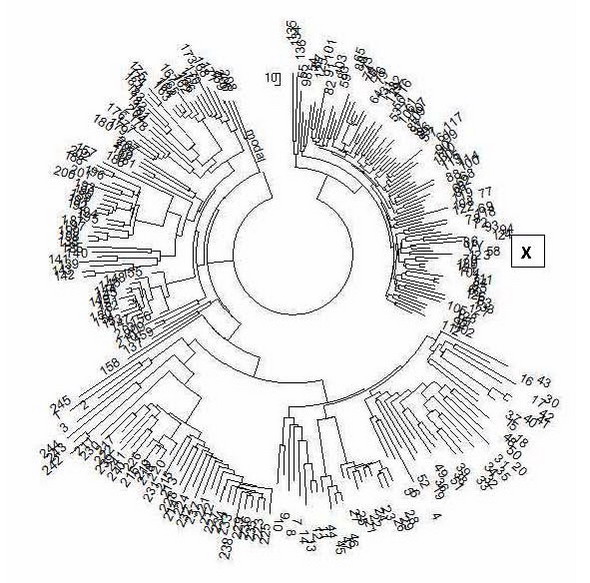













| Lasīt: |
|---|

Jauns
- Altaja valodu saimē ietilpst
- Gruzīnu valodas un tūrisma vārdnīca
- Ir tikai strukturālās un objektu programmēšanas paradigmas Programmēšanas paradigmas un tehnoloģijas
- Armēņu alfabēts kā ķīmisko elementu periodiskās tabulas kods
- Slāvi un substrāts (fragments no pārskatītās versijas par haplogrupu I2a1)
- "Fets Afanasijs Afanasjevičs
- Ģeodēziskie darbi lineāriem objektiem
- Balmonta dzejoļa “Es nācu šajā pasaulē...
- Gatavs elektroniskais portfolio bērnudārza audzinātājai
- Jauniešu satiksmes drošības brigādes runa pamatskolā. Jauniešu brigādes loma skolā




