Сайтын хэсгүүд
Редакторын сонголт:
- Мэдээллийн технологийн ажилтнууд нь инженер-техникийн ажилчид (хичээл)
- Ял шийтгүүлсэн хүнийг нийгэмшүүлэх 14 үндсэн чиглэл
- Вячеслав Наговицын ажил олдсонгүй
- Алексей Пушков: намтар, хувийн амьдрал, гэр бүл, гэрэл зураг Хүүхэд нас, гэр бүл
- Печерскийн Гэгээн Сисой, схем лам Печерскийн Ариун Эрхэм лам Иосаф
- Есүс Христийн амилалт Христийн амилалтын дараа юу болсон
- Ариун онгон Мариагийн сүм Ариун онгон Мариагийн сүм
- Ортодокс нэвтэрхий толь бичгийн мод дахь Дөрөв дэх Менионы утга
- Эрүүл мэндийн яамны 02-ны өдрийн 290н тоот тушаал
- Экспоненциал тэгш бус байдал
Зар сурталчилгаа
| I2 гаплогрупп. Славян ба субстрат (I2a1 гаплогруппын талаархи шинэчилсэн хувилбарын хэсэг) |
|
111 маркерын гаплотипийг хамгийн нарийвчилсан байдлаар авсан бөгөөд тэдгээрийн дотор танилцуулсан гаплотиптэй яг адилхан эхний 12 маркер бүхий 15 гаплотип байдаг нь тогтоогджээ. Эдгээр нь I1 хаплогруппын 968 111 маркер гаплотипийн жагсаалтын дагуу 57-60, 110, 131, 132, 153-157, 660, 744, 745 дугаартай гаплотипууд юм. Дараа нь мэргэжлийн компьютерийн программ (PHYLIP) ашиглан бүх 968 111 маркерын гаплотипээс модыг барьсан бөгөөд хамгийн холбоотой (эхний 12 маркерын дагуу) гаплотипуудыг тодорхойлсон. Үүссэн модыг тайлбарлах зорилгоор доор үзүүлэв. Энэ нь сайн тэгш хэмийг харуулдаг бөгөөд энэ нь мод бүхэлдээ нэг нийтлэг өвөг дээдсээс гаралтай болохыг харуулж байна. Түүний хэзээ амьдарч байсныг бас харуулах болно. Мэдээллийн хувьд бүхэл бүтэн модны үндсэн (өвөг дээдсийн) гаплотип нь дараах байдалтай байна 13 22 14 10 13 14 11 14 11 12 11 28 – 15 8 9 8 11 23 16 20 28 12 14 15 16 – 10 10 19 21 14 14 16 20 35 37 12 10 – 11 8 15 15 8 11 10 8 9 9 12 23 25 15 10 12 12 16 8 13 25 20 13 13 11 12 11 11 12 11 – 32 12 8 17 12 24 27 19 11 12 12 13 11 9 11 11 10 12 12 31 11 13 21 16 11 10 24 15 19 11 24 17 13 15 25 12 22 18 12 14 18 9 12 11 Эхний 12 маркерт үзүүлсэн гаплотип нь үүнээс зөвхөн нэг мутациар ялгаатай (дараалсан тав дахь мутаци, тодруулсан). Нийтдээ бүх 968 гаплотип нь эхний 67 маркерын самбараас 14,891 мутаци, бүх 111 маркер дээр 24,990 мутацийг харуулж байна. Энэ нь нийтлэг өвөг дээдэс нь 14891/968/0.12 = 128 → 147 уламжлалт үе (тус бүр 25 жил), эсвэл 24990/968/0.198 = 130 → 149 уламжлалт үе (0.12 ба 0.198 нь 1-ийн мутацын хурд, 17-ын тогтмол 17-) амьдарч байсныг харуулж байна. маркер гаплотипууд, сум – давтагдах мутацийн хүснэгтийн засвар), эсвэл нийтлэг өвөг дээдсээс 3675±370 ба 3725±375 жилийн өмнө тус тус. Эндээс харахад 67 ба 111 маркерын гаплотипууд бараг ижил үр дүнг өгдөг бөгөөд 3700 жилийн дэвсгэр дээр ердөө 50 жилийн зөрүүтэй байдаг.
Ижил 12 маркер/аллелийг агуулсан эдгээр долоон гаплотип нь модны гурван өөр мөчир дээр байрладаг нь тогтоогджээ. Мэдээжийн хэрэг, хэрэв танилцуулсан гаплотип нь 25, 37 ба түүнээс дээш тэмдэглэгээтэй байсан бол зөвхөн нэг мөчир гарч ирэх нь гарцаагүй, гэхдээ энэ нь тийм юм. Зарчмын хувьд энэ нь бидэнд нэг их саад болохгүй. Эдгээр салбарууд нь дараах байдалтай байна.
13 22 14 10 13
14 11 14 11 12 11 28 — нийтлэг өвөг дээдсээс 1600±240 жилийн өмнө (Танилцуулсан гаплотипийн гурван мутацийг тодруулсан). Үзүүлсэн гаплотип нь зарчмын хувьд эдгээр салбаруудын аль нэгэнд байж болно; түүний зөвхөн 12 тэмдэгт нь дөрвөн салбараас аль нэгийг нь хасах боломжийг бидэнд олгодоггүй. Эдгээр дөрвөн салааны нийтлэг өвөг дээдэс нь өвөг дээдсийн гаплотипуудын баруун талд амьдардаг байв. Дээрх тоонуудтай холбоотой гаплотипууд аль дэд бүлэгт хамаарах, түүний эхний 12 тэмдэглэгээ бүрэн давтагдаж байгааг харцгаая. Энэ материалын төгсгөлд эдгээр хүмүүсийн дөрвийн геномын мэдээллийг өгсөн бөгөөд үүнд M253-ийн доод талын SNP-ийг тэмдэглэсэн болно. Хамгийн магадлалтай, ID00020 нь ижил зүсэлттэй байх болно. Ерөнхийдөө эдгээр гаплотипуудын дийлэнх нь SNP-тэй: I1-M253 > DF29 > CTS6364 > M227, өөр салбарт I1-M253 > DF29 > Z58 > Z138. ID00020 snips нь эхний эсвэл хоёр дахь гинжин хэлхээнд хамаарах ёстой. Эдгээрийг бүгдийг нь Москвагийн ДНХ-ийн удмын сангийн лабораторид тогтооно. Үзүүлсэн гаплотип хүртэлх түүхэн зам нь ойролцоогоор дараах байдалтай байна. Хаплогрупп I нь 43 мянган жилийн өмнө (IJ хосолсон гаплогруппээс) үүссэн бөгөөд 28 мянган жилийн өмнө I1 ба I2 гаплогруппууд болон хуваагдсан (хэдийгээр "ялгасан" гэдэг нь харьцангуй нэр томъёо боловч тэд салаагүй, эдгээр нь бие даасан үйл явдлууд байсан) , эхнийхээс (хоёр дахь шиг) олон дэд бүлгүүд цаг хугацааны явцад үүссэн. Хаплогрупп I нь Швед болон Төв Европт 7000 жилийн тэртээ олдсон чулуужсан араг ясны үлдэгдлээс олдсон. 4600-4000 жилийн өмнө Эрбинс Европыг суурьшуулах үеэр Төв Европоос хаплогрупп I1, R1b хаплогрупп тээгч нар бараг бүрэн алга болсон ч хэн нэгэн амьд үлдсэн бөгөөд үүний үр дүнд I1 гаплогруппын орчин үеийн тээвэрлэгчид харьцангуй Ойролцоогоор 3700 жилийн өмнө, өөрөөр хэлбэл МЭӨ 2-р мянганы эхний хагаст амьд үлдсэн үр удмыг бий болгосон сүүлийн үеийн нийтлэг өвөг дээдэс. Гаплогрупп I1-д зөвхөн нэг нийтлэг өвөг байдаг тул гаплотипийн мод нь тэгш хэмтэй байдаг. Энэ нь маш ховор тохиолддог. ID00020 нь энэхүү нийтлэг өвөг дээдсийн удам юм. Өргөдөл. Энд бид ID00020 салбараас авсан гаплотип тээгчдээс олдсон SNP-ийн найрлагын талаарх дэлгэрэнгүй геномын өгөгдлийг өгдөг. Энэ нь терминалын snips-ийг эс тооцвол бараг ижил байх ёстой. Гэхдээ дээр дурдсан хэд хэдэн тохиолдлуудын адил эдгээр нь адилхан байж магадгүй юм. M223 болон доорх SNP-уудыг онцлон тэмдэглэв: Иж бүрдэл дугаар 38184 (Мэтью Хамилтон Батчер, Америкийн индианчууд, гэхдээ дээрх салбар дээрх 110 гаплотипийн европоос Y-ДНХ авсан): DF29+, M227+, CTS10058+, CTS10140+, CTS10338+, CTS10834+, CTS11036+, CTS11042+, CTS11441+, CTS11526+, CTS11552+, CTS117152+, CTS117152+, CTS117155, CTS3+ CTS2193+, CTS2375+, CTS2514+, CTS2524+, CTS2644+, CTS2738+, CTS3517+, CTS3654+, CTS3843+, CTS4088+, CTS4130+ , CTS4295+, CTS4437+, CTS4848+, CTS4982+, CTS5167+, CTS5408+, CTS5513+, CTS565+, CTS5650+, CTS5705+, CTS571+, CTS5783+, CTS57+, CTS58+, CTS5+, CTS59 93+, CTS6140+, CTS6221+, CTS623+, CTS6265+, CTS6395+, CTS641+, CTS6629+, CTS674+ , CTS6932+, CTS7267+, CTS7329+, CTS7831+, CTS7949+, CTS8333+, CTS8345+, CTS8394+, CTS8420+, CTS8716+, CTS88+, CTS8824+, CTS8824+, CTS8824+, CTS8824+, CTS8824+, 09+, F3692 +, F719+, L118+, L121+, L123+, L124+, L125+, L132+, L15+, L16+, L187+, L350+, L403+, L41+, L468+, L470+, L498+, L509+, L574+, L575+, L578+, L740+, L748+, L75+, L75+, L75+, L75+, L75, L75+ 759+, L772 +, L80+, L81+ , M168+, M170+, M213+, M227+, M235+, M253+, M258+, M294+, M299+, M307+, M89+, M94+, P123+, P124+, P125+, P126+, P127+, P129+, P130+, P133+, P134+, P136+, P14+, P13+, P14+, P13+, P14+, P14+, P14+ , P141+, P143+, P145+, P146+ , P148+, P149+, P151+, P157+, P158+, P159+, P160+, P161+, P163+, P166+, P187+, P212+, P30+, P316+, ХУУДАС00026+, PAGES, PAGES00026+0, PAGES, PAGES+ F2608+, PF2611+, PF2615+, PF2747+, PF2748+, PF2749+, PF2770+ , PF3562+, PF3574+, PF3639+, PF3640+, PF3641+, PF3660+, PF3666+, PF3672+, PF3675+, PF3677+, PF3780+, PF3800, P380F, PF3800, P380, P3+ 3814+, PF3815+, PF3819+, PF3822+, PF3836+, PF3837+, PF6464+, PF6469+, PF6470+ , PF6477+ , PF6479+, PF6520+, V241+, V250+, YSC0000207+, YSC0000227+, YSC0000256+, YSC0000272+, YSC0000281+, YSC0000281+, YSC1, M34, M30+, M39, M30+ M72-, Z58-, Z63-, Z17694-, S2348-, S4442-, M3453- Иж бүрдэл дугаар 38737 (Greathouse, Герман, гаплотип 744): CTS10058+, CTS10140+, CTS10338+, CTS10362+, CTS109+, CTS11036+, CTS11042+, CTS11358+, CTS1114+, CTS1114+, CTS5114+ 5 75+, CTS11726+, CTS11775+, CTS11783+, CTS11950+, CTS125+, CTS12632+, CTS1393+, CTS1996+, CTS2193+, CTS2375+, CTS2514+, CTS2524+, CTS2536+, CTS2644+, CTS2738+, CTS3331+, CTS3431+, CTS3517+, CTS3536+, C36+, CTS36+, C64+, CTS38+ 68+, CTS39 96+, CTS4088+, CTS4130+, CTS4295+, CTS4364+, CTS4368+, CTS4443+, CTS4740+, CTS4848+, CTS4982+, CTS5167+, CTS5318+, CTS5408+, CTS5457+, CTS5513+, CTS5532+, CTS565+, CTS5650+, CTS5705+, CTS571+, CTS5708+, CTS571+, CTS598+, CTS5985 +, CTS6135+, CTS6140 +, CTS6221+, CTS6265+, CTS6383+, CTS6395+, CTS641+, CTS6629+, CTS674+ , CTS6800+, CTS6907+, CTS6932+, CTS7267+, CTS7329+, CTS7502+, CTS7831+, CTS7922+, CTS7933+, CTS7949+, CTS8243+, CTS8333+, CTS8334+, CTS833+, CTS84+ 8716+, CTS88+, CTS8876+, CTS8980+, CTS9240+, CTS9264+, CTS9288+, CTS9828+, F1046+, F1209+, F1302+, F1320+, F1329+, F1450+, F1460+, F1704+, F1714+, F1753+, F1767+, F2048+, F2075+, F2142+, F21502, F36+, F21502, F21502, F36+ +, F2408+, F2587+, F2688 +, F2710+, F2794+, F2837+, F2985+ , F2993+, F3033+, F3111+, F3136+, F3335+, F3368+, F3402+, F3556+, F3692+, F4188+, F719+, F922+, L118+, L121+, L12+, L12+, L12+, L15, L15 , L16+, L187+, L350+, L403+, L468+, L470+ , L498+, L509+, L574+, L575+, L578+, L740+, L748+, L75+, L750+, L751+, L755+, L756+, L758+, L759+, L772+, L80+, M17+, M17+, L80+, M17+, M18+ M253+ Z138+, Z139+, M170+, M258+, M307+, P19+, P30+, P38+, P109-, P259-, P37-, M161-, M72-, M26-, M21-, M223-, M227- Иж бүрдэл дугаар 152077 (Jens Peter Nielsen, Дани, гаплотип 745): CTS10058+, CTS10140+, CTS10338+, CTS10362+, CTS109+, CTS11036+, CTS11042+, CTS11358+, CTS11415+, CTS11358+, CTS1415+, C TS11575+, CTS11726+, CTS11775+, CTS11783+, CTS11950+, CTS125+, CTS12632+, CTS1393+ , CTS1996+, CTS2193+, CTS2375+, CTS2514+, CTS2524+, CTS2536+, CTS2644+, CTS2738+, CTS3331+, CTS3431+, CTS3517+, CTS3536+, CTS36+, CTS36+, CTS36+, CTS36+, CTS36+ 38 68+, CTS3996+, CTS4088+, CTS4130+, CTS4295+, CTS4364+, CTS4368+, CTS4443+, CTS4740+ , CTS4848+, CTS4982+, CTS5167+, CTS5318+, CTS5408+, CTS5457+, CTS5513+, CTS5532+, CTS565+, CTS5650+, CTS5705+, CTS571+, CTS58+, CTS58+, CTS58+, CTS589 93+, CTS6135+, CTS6140+, CTS6221+, CTS6265+, CTS6383+, CTS6395+, CTS641+, CTS6629+, CTS674+, CTS6800+, CTS6907+, CTS6932+, CTS7267+, CTS7329+, CTS7502+, CTS7831+, CTS7922+, CTS7933+, CTS7949+, CTS8243+, C333+, CTS4+8, C33+, CTS8+ 0+, CTS8716+, CTS88 +, CTS8876+, CTS8980+, CTS9240+, CTS9264+, CTS9288+, CTS9828+, F1046+ , F1209+, F1302+, F1320+, F1329+, F1450+, F1460+, F1704+, F1714+, F1753+, F1767+, F2048+, F2075+, F2142+, F263+, F2142+, F2625+, F2625+ 02+, F2408+, F2587 +, F2688+, F2710+, F2794+, F2837+, F2985+, F2993+, F3033+, F3111+, F3136+, F3335+, F3368+, F3402+, F3556+, F3692+, F4188+, F719+, F922+, L118+, L15+, L118+, L12+, L12+, L12+ L157+, L16+, L187+, L350+, L403+, L468+ , L470+, L498+, L509+, L574+, L575+, L578+, L740+, L748+, L75+, L750+, L751+, L755+, L756+, L758+, L759+, L7072, M18+, L759+, L7072, M18+, M18+, M18+ +, M253+, M294+, M307+, M42+, M450+, M89+, M94+, P123+, P124+, P126+, P127+, P130+, P135+, P136+, P138+, P14+, P141+, P14+, P14+, P15+, P115+ P159+, P160+, P166+, P187+ , P30+, P305+, PAGES00026+, PAGES00081+, PAGES00123+, PF1016+, PF1029+, PF1031+, PF1040+, PF1046+, PF1061+, PF109+, PF1061+, PF109+, P109F, P17F, P+19F 1269+, PF1276+, PF192+, PF210+, PF212+, PF223+, PF234+, PF258+, PF2591+ , PF2593+, PF2599+, PF2608+, PF2611+, PF2615+, PF2624+, PF263+, PF2643+, PF272+, PF2745+, PF2747+, PF2748+, PF2748+, P274+, P27F, P7F29, 2+, PF316+, PF325+, PF342+, PF3515+, PF3517+, PF3518+, PF3534+, PF3560+, PF3561+ , PF3562+, PF3574+, PF3578+, PF3586+, PF3588+, PF3590+, PF3594+, PF3596+, PF3600+, PF3604+, PF3605+, PF3611F, PF3611F, P3F61F, P3611F F3625+, PF 3639+, PF3640+, PF3641+, PF3642+, PF3654+, PF3660+, PF3666+, PF3672+ , PF3675+, PF3677+, PF3686+, PF3694+, PF3780+, PF3800+, PF3804+, PF3806+, PF3807+, PF3809+, PF3811+, PF3814+, PF3814, P38+, P3814, P381F F3836+, PF3837+, PF 500+, PF667+, PF719+, PF725+, PF779+, PF796+, PF803+ , PF815+, PF821+, PF840+, PF844+, PF892+, PF937+, PF951+, PF954+, PF970+, V168+, V186+, V189+, V205+, V221+, V, SC+, V21+, V, 02, V, 02 56+, YSC0000256+, YSC00002 57+, YSC0000259+, YSC0000260+, YSC0000264+ , YSC0000265+, YSC0000267+, YSC0000272+, YSC0000280+, YSC0000281+, YSC0000298+, YSC0000299+, YSC0000300+, YSC0000300+, Z138+, Z139+, M170+, M258+, M307+, P19+, P30+, P38+, P109-, P259-, P37-, M161-, M72-, M26-, M21-, M223-, M227- Kit N10060 (Роберт Уэйд, Англи, гаплотип 57): M170+, M253+, M258+, M307+, P19+, P30+, P38+, P109-, P259-, P37-, M161-, M72-, M26-, M21-, M223-, M227- Haplogroup I2a Хоёр хүн ийм хаплогрупптэй байсан бөгөөд нэг нь I2a1-L147.2 дэд бүлэг, хоёр дахь нь - I2a2-M223, 67 ба 111 маркер гаплотиптэй. KLIN ID00004 67 маркерын гаплотипийг танилцуулж, I2a хаплогруппийг мэдээлэв. 13 24 17 11 14 15 11 13 12 13 11 30 – 17 8 10 11 11 25 15 20 0 12 14 15 15 – 10 10 20 21 15 12 19 17 35 35 11 10 – 11 8 15 15 7 12 10 8 11 9 12 22 22 16 10 12 12 12 7 10 30 21 13 14 10 13 11 11 12 9 Энэхүү гаплогрупп нь Зүүн Европ даяар тархсан байдаг, тухайлбал Грек, Серби, Босни-Герцеговина, Македон, Чех, Словак, Польш, Болгар, Беларусь, Орос, Украин, Унгар, түүнчлэн Герман, Румын, бага зэрэг. Итали болон Туршилтанд хамрагдсан хүмүүсийн дийлэнх нь L147.2 гэгддэг I2a-CTS10228 SNP-тэй болох нь тогтоогдсон (доорх ISOGG диаграммыг үзнэ үү; баруун талд YFull компанийн дагуу SNP үүсэх он сар өдөр байна). Энэ бол Зүүн Европын ердийн snip юм. Гаплотипийн дүр төрхийг харахад үйлчлүүлэгчид ч бас байгаа нь тодорхой байна. Асуулт нь: бид гаплотипийн төрлөөс хамааран доод түвшний SNP-ийг урьдчилан таамаглаж чадах уу?
Мэдээллийн сан (FTDNA I2a Төсөл) нь 67 тэмдэглэгээний форматтай 245 гаплотип агуулдаг.Эдгээр бүх гаплотипуудаас бүтээгдсэн модны баруун талд ID00004 гаплотип (X тэмдэглэгдсэн) хамгийн их эзэлдэг холбогдох томоохон салбаруудад харьяалагддаг. баруун, авсаархан мөчир модны.Авсаархан гэдэг нь сул мөчрүүдтэй харьцуулахад харьцангуй залуу мөчир гэсэн үг.Өөрөөр хэлбэл авсаархан мөчир нь харьцангуй сүүлийн үеийн гарал үүслийг илэрхийлдэг.Тооцооллоос харахад энэ мөчир ойролцоогоор 2300 жилийн настай, өөрөөр хэлбэл үүссэн. Өнгөрсөн эриний төгсгөлд. Энэ салбарын бараг бүх гаплотипууд нь CTS10228/L147.2 snip-тэй холбоотой байдаг ч жишээлбэл, уруудах snips байдаг. L147.2 >YP204/S17250 >Z16971 Эдгээр нь мод дээрх 67 маркерын гаплотипоор тусгаарлагдаагүй. Илүү нарийвчлалтай хэлбэл, тэдгээр нь салбарын өөр өөр хэсгүүдэд хуваагдаж болох ч Төслийн хэт цөөн тооны гаплотип тээгч ийм гүн гүнзгий SNP-ийг тодорхойлсон байдаг. Тэднийг Москвагийн ДНХ-ийн удмын сангийн лаборатори тогтооно.
111 тэмдэгт мод дээр тодорхой зүсэлттэй холбоотой тодорхой мөчрүүд байдаггүйг бид харж байна. Салбарын үндсэн гаплотип 67 маркер формат: 13 24 16 11 14 15 11 13 13 13 11 31 – 17 8 10 11 11 25 15 20 32 12 14 15 15 – 10 10 21 21 15 12 18 18 34 35 11 10 – 11 8 15 15 7 12 10 8 11 9 12 22 22 16 10 12 12 12 7 10 30 21 13 14 10 13 11 11 12 9 (Танилцуулсан гаплотипээс 8 мутацийг тодорхойлсон). Энэ нь гаплотип нь өвөг дээдсээс 8 / 0.12 = 67 → 72 уламжлалт үе буюу ойролцоогоор 1800 жилээр хасагдсан гэсэн үг юм. Энэ нь мэдээжийн хэрэг ойролцоогоор юм, учир нь хоёр гаплотипийн яг нарийн тооцоо байхгүй - танилцуулсан гаплотип дахь санамсаргүй нэг мутаци нь тооцоолсон зайг 200 гаруй жилээр солих болно. Ямар ч байсан захиалагчийн өвөг дээдэс ID00004 ДНХ-ийн шугамыг энэ салбарын нийтлэг өвөг дээдсээс бараг шууд олж авсан нь тодорхой. Ойролцоогоор 2300 жилийн настай I2a-L147.2 хаплогрупп/ дэд бүлгийн түүхийг Reformat дээр олон удаа хэлэлцсэн. Хаплогрупп I2a нь 7000-5000 жилийн өмнөх үетэй төв Европ дахь оршуулгын газраас олдсон бөгөөд дараа нь 4500 орчим жилийн өмнө Төв Европоос алга болж, хоёр салаа болж хуваагдсан. Нэг (илүү нарийвчлалтай, хэд хэдэн салбар, дээрх эхний диаграмыг харна уу) Британийн арлууд руу нүүсэн (эсвэл зугтсан) бөгөөд энэ салбаруудын нийтлэг өвөг дээдэс нь ойролцоогоор 4500-5000 жилийн өмнөх үеэс эхэлдэг. I2a-L147.2 дэд бүлгийн өөр нэг салбар Европт хүн амын түгжрэлийг даван туулж, 2300 орчим жилийн өмнө Дунай-Карпатын нуруунд сэргэж, МЭ 1-р мянганы эхний хагаст Зүүн Европ даяар тархсан. Түүхчид, археологичид үүнийг "Славуудын гарал үүсэл" гэж андуурсан байж магадгүй бөгөөд энэ нь "Өнгөрсөн он жилүүдийн үлгэр" зэрэг эртний түүхүүдэд энэ тухай яригддаг. Хэрэв PVL зөв бол I2a-L147.2 гаплогруппын славянчууд Адриатын тэнгисийн хойд хэсэгт орших Альпийн нурууны зүүн хэсэгт орших Норикоос ирсэн. Норикийн оршин суугчид болох Таурисканчууд МЭӨ 1-р мянганы хоёрдугаар хагасаас хойш мэдэгдэж байсан. Хэрэв ID00004 нь BigY тест (эсвэл Y хромосомын өөр геномын сорил) авсан бол үр дүн нь дараахтай ойролцоо (эсвэл ижил) байх байсан (I2a-CTS10228/L147.2 дэд бүлгийн өндөр SNP-ийг энд онцолсон болно). Үлдсэн тэмдэглэгдсэн SNP нь илүү өндөр гаплотипуудад хамаардаг, жишээлбэл, M42 - хосолсон гаплогрупп VT; V9, V52 ба V189 бүгд хосолсон ST haplogroup; M89 нь F гаплогрупп, L15 ба L16 нь IJK гаплогрупп, L41 нь I гаплогрупп гэх мэт. Таны харж байгаагаар BigY тестийн (599 доллараар) өгсөн цорын ганц ашигтай зүйл бол хамгийн бага I2a-CTS10228 snip-ийг тодорхойлох явдал байв. Энд дахиж дагалдах snip байхгүй бөгөөд доорх жагсаалтын дээрх Z17855 snip сөрөг байна (эцсийн дөрөв дэх). Доор өгөгдсөн зургийн багцыг нээлттэй мэдээллийн сангаас авсан бөгөөд энэ нь Польшийн хунтайж Святопольк-Четвертинскийн удамд харьяалагддаг бөгөөд домог ёсоор Польшийн Руриковичуудаас гаралтай. Дашрамд дурдахад, Наполеоны дайны оролцогч, Оросын армийн хурандаа хунтайж Борис Антонович Четвертинский (1784-1865) нь Польшийн нэгэн гэр бүлээс гаралтай. Бүгд биш юмаа гэхэд ихэнх хэсэг нь ID00004-с олдох ёстой. Гэсэн хэдий ч, энэ нь зөвхөн гүн дэд давхарга нь Четвертинскийнхтэй яг адилхан болсон тохиолдолд л болно. Гэхдээ Chetvertinsky-д L147.2-оос доош гүн зүсэлт олдоогүй нь сонирхолтой юм, хамгийн сүүлчийн терминал нь CTS10228, өөрөөр хэлбэл L147.2 юм. CTS4002+, CTS4039+, CTS4088+, CTS410+, CTS4437+, CTS4848+, CTS4982+, CTS5044+, CTS5375+, CTS5650+, CTS5727+, CTS5884+, C69+, CTS5884+, C69+, C69+ +, CTS6265+, CTS674+, CTS6932+, CTS7175+, CTS7213+, CTS7218+, CTS7329+, CTS7831+, CTS8239+ , CTS8333+, CTS8345+, CTS8420+, CTS8486+, CTS88+, CTS8876+, CTS9240+, CTS9264+, CTS9349+, CTS10058+, CTS10228+, CTS10834+, CTS10936+, CTS11030+, CTS11441+, CTS11768+, CTS1293+, CTS176+, CTS1846+, CTS2193+, CTS2514+, CTS3296+, CTS2514+, CTS3296+, CTS177, C403+, CTS195+ F3692+, F719+, L132+, L15+, L16+, L178+, L350+, L403+, L41+, L460+, L468+, L470+, L498+, L578+, M42+, M423+, M438+, M89+, M94+, P123+, P124+, P125+, P126+, P127+, P129+, P130+, P133+, P134+, P135+, P136+, P138+, P139+, P14+, P14+, P14+, P14+, P14+, P14+, P14+ , P146+, P148+, P149+, P151+ , P157+, P158+, P159+, P160+, P161+, P163+, P166+, P187+, P212+, P316+, P38+, PAGES00026+, PAGES00081+, PF2591+, P67+, PF160, P64F, P6+F 7+, PF2748+, PF2749+, PF2770+, PF3562+, PF3574+, PF3626+ , PF3638+, PF3639+, PF3640+, PF3641+, PF3644+, PF3651+, PF3652+, PF3657+, PF3658+, PF3660+, PF3664+, PF366+, PF366, P3F66+, PF366, P3F66+ F3672+, PF 3675+, PF3677+, PF3780+, PF3800+, PF3801+, PF3802+, PF3804+, PF3809+ , PF3811+, PF3812+, PF3814+, PF3815+, PF3819+, PF3820+, PF3822+, PF3836+, PF3837+, PF3849+, PF3876+, PF396+, P646, P6+, P6+, P64+ F6470+, PF6477+, PF 6479+, PF6520+, L621+, L68+, L748+, L751+, L755+, L756+, L758+, L772+, M168+, M170+, M213+, M235+, M258+, M294+, M299+, V241+, V250+, YSC0000207+, YSC0000227+, YSC0000256+, YSC0000272+, M423+, V23, Z712, S712- -, Z2109-, M3453-, S2348-, S4442- , P61-, L584-, CTS3802-, DF41- Бид Руриковичийн тухай ярьж байгаа тул Оросын язгууртны чуулганы ноёдын бүлэг N1c1 гаплогрупптэй бол Четвертинский (болон үйлчлүүлэгч) I2a гаплогрупптэй гэдгийг санацгаая. Рурик, хэрэв ийм зүйл байсан бол зөвхөн нэг гаплогрупптэй байж болно. Тиймээс олон тооны Рюриковичийн тухай асуулт, хэрэв тэд бүгд Рурикович гэж нотлох баримтат үндэслэлтэй бол нээлттэй хэвээр байна. Миний мэдэж байгаагаар хэнд ч баримтат нотлох баримт байхгүй, гэхдээ ноёдын угийн бичгийн талаар шинжээчдийн баттай дүгнэлт байдаг. Тэгээд ч Польшийн угийн бичигчид нэг бодолтой байхад Оросын удам судрууд өөр бодолтой байдаг байх. Тэд өөр хоорондоо юмыг олох гэж оролдож байж болох ч ДНХ-ийн удмын бичиг нь тэдэнд авч үзэх шинэ мэдээллийг өгдөг. Магадгүй Москвагийн ДНХ-ийн удмын сангийн лаборатори ID00004-ийн доод талын SNP-ийг (CTS1028-тай холбоотой) тодорхойлж, аль бүс нутагт голчлон илэрхийлэгдэж байгааг ойлгох боломжтой байх. Тэгвэл хувийн угийн бичгийг улам гүнзгийрүүлж болно. KLIN ID00019 111 маркерын гаплотипийг танилцуулсан бөгөөд I2b хаплогрупп, I2b1 дэд бүлэг, M223 секц гэж мэдээлсэн. 15 23 15 10 15 15 11 14 12 14 12 32 16 8 9 11 11 25 14 20 28 11 14 14 15 11 10 19 21 15 14 18 21 32 40 12 10 11 8 16 16 8 11 10 8 10 9 12 22 22 15 11 12 12 15 9 14 27 21 11 13 12 12 11 12 12 11 32 13 8 15 10 24 27 16 12 11 14 11 12 9 13 11 10 10 12 32 10 12 22 13 11 10 20 15 23 9 24 14 12 14 27 12 21 18 12 15 17 9 12 11 Үзүүлсэн өгөгдөл нь 2010 оноос хойш хоцрогдсон нэр томъёог ашигласан бөгөөд үүнээс хойш SNP M223 нь I2a2a дэд ангилалд багтсан (доорх диаграмыг товчилсон хэлбэрээр харна уу). Баруун талын баганад YFull компанийн геномын өгөгдлийг ашиглан тооцоолсон хамгийн эртний болон бусад зарим SNP-ийн гарал үүслийн ойролцоогоор он сар өдрийг (олон жилийн өмнө) харуулав.
Дээрх голт бор өнгө нь I2a-L147.2 дэд бүлгийг харуулж байгаа бөгөөд энэ нь Зүүн Европ дахь I haplogroup-д хамгийн түгээмэл байдаг - Грекээс Балтийн орнууд, түүний дотор Орос, Украин, Беларусь, хуучин Югославын орнууд; Улаан шугам нь танилцуулсан гаплотипийн дэд бүлэг болох I2a-M223-ийг онцолж өгдөг. Дэд бүлэг нь өөрөө эртний бөгөөд 17,400 жилийн настай бөгөөд мэдээжийн хэрэг, энэ нь доод дэд бүлгүүдтэй бөгөөд бид энэ тайлбарт үүнийг тодорхойлоход ойртох болно. ID00019-д байж болох дэд бүлгүүдийг доор харуулсны дагуу шараар тодруулсан болно. FTDNA I төслийн I2 гаплогруппын 111 маркер гаплотипийн ерөнхий модон дээрх танилцуулсан гаплотипийн байршлыг тодорхойлохыг хичээцгээе. Үүнийг хийхийн тулд бид төслийн I2 гаплотипийн бүх 63 111 маркер гаплотипийг авч, тэдэнд танилцуулсан гаплотип ID00019-ийг нэмж, мэргэжлийн компьютерийн программ (PHYLIP) ашиглан үүссэн 64 гаплотипээс мод бүтээх болно. Энэхүү туршилтын зорилго нь танилцуулсан гаплотипийг аль гаплотип, ямар дэд бүлгээр хүрээлэхийг олж мэдэх явдал юм. I2 хаплогруппын эдгээр 111 маркер гаплотипуудын модыг доор өгөв.
Үзүүлсэн гаплотипийн байрлалыг X индексээр модон дээр зааж өгсөн болно. Энэ нь гадаад төрхөөрөө ижил төстэй, магадгүй холбоотой гаплотипуудаас бүрдсэн тусдаа, сайн тодорхойлогдсон салбар юм. Мэдээллийн хувьд I2 модны бүхэл бүтэн үндсэн (өвөг дээдсийн) гаплотип нь дараах байдалтай байна 13 24 16 10 13 15 11 13 12 13 11 30 17 8 10 11 11 25 15 20 29 12 14 15 15 10 10 19 20 15 14 17 18 34 36 12 10 11 8 15 16 8 11 10 8 11 9 11 21 22 16 11 12 12 15 8 13 25 21 11 13 11 13 11 12 12 11 30 14 8 15 11 25 27 18 12 11 12 12 12 9 12 11 10 11 12 31 11 12 22 14 11 10 22 15 20 11 23 16 11 15 25 12 22 18 12 14 17 9 12 11 Үзүүлсэн гаплотип нь үүнээс 70 мутациар ялгаатай. Энэ нь 70/0.198 = 354 → 537 ердийн үеийнхтэй тэнцүү буюу тэдгээрийн хооронд ойролцоогоор 13,425 жил байна (0.198 нь 111 маркерын гаплотипуудын мутацийн хурдны тогтмол, сум нь давтагдах мутацийн залруулга). Өөрөөр хэлбэл, хаплогруппын нийтлэг өвөг нь ID00019-ээс цаг хугацааны хувьд маш хол байдаг. Бүх 64 гаплотип нь үндсэн гаплотипээс 3906 мутацитай бөгөөд энэ нь энэ гаплотипийн орчин үеийн тээгчдийн нийтлэг өвөг дээдэс амьдарч байсан үетэй тохирч байгаа бөгөөд энэ тохиолдолд 3906/64/0.198 = 308 → 440 нөхцөлт үе (тус бүр 25 жил, Энэ хугацаанд тэдгээрийг тохируулсан мутацийн хурдны тогтмолууд), эсвэл 11000±1100 жилийн өмнө. Тэрээр I2 дэд бүлэг (ойролцоогоор 27,500 жилийн өмнө, YFull-ийн хэллэгээр тодорхойлогдсон) үүссэнээс хойш нэлээд удаан амьдарч байсан нь тодорхой бөгөөд энэ нь I2 хаплогрупп нь 11,000 жилийн өмнө Европ дахь мөстлөгийн үетэй холбоотой байж болзошгүй хүн амын хүнд хэцүү хүндрэлийг даван туулж байсныг харуулж байна. . Үйлчлүүлэгч ба I2 гаплогруппын нийтлэг өвөг дээдэс нь ойролцоогоор (13425+11000)/2 = 12200 жилийн өмнө, өөрөөр хэлбэл гацаа үүсэхээс өмнө амьдарч байжээ. Үзүүлсэн гаплотип нь 39, 44-46, 51, 52 дугаартай гаплотипуудтай нэг салбар дээр байгааг бид харж байна. Энэ салааны үндсэн гаплотип. 15 23 15 10 15 15 11 13 12 14 12 32 15 8 10 11 11 25 14 20 28 11 14 14 15 11 10 19 21 14 14 17 19 33 39 12 10 11 8 15 16 8 11 10 8 10 9 12 21 22 15 11 12 12 14 9 13 27 20 11 13 12 12 11 12 12 11 31 13 8 15 11 24 27 16 12 11 13 11 13 9 12 11 10 11 12 32 10 12 22 13 11 10 20 15 22 9 23 14 12 14 28 12 21 18 12 15 17 9 12 11 Энэ нь танилцуулсан гаплотипээс 23 мутаци (тэмдэглэгдсэн) -ээр ялгаатай, өөрөөр хэлбэл гаплотип нь энэ салбар дахь өвөг дээдсээс 23/0.198 = 116 → 132 ердийн үе (тус бүр 25 жил), өөрөөр хэлбэл ойролцоогоор 3300 жилээр хасагдсан байна. Үүний зэрэгцээ, салбарын бүх долоон гаплотип нь дээр үзүүлсэн үндсэн гаплотипээс 163 мутацитай, өөрөөр хэлбэл энэ салааны нийтлэг өвөг 163/7/0.198 = 118 → 134 ердийн үе, өөрөөр хэлбэл 3350 ± 425 жил амьдарч байжээ. өмнө. Эдгээр хоёр утга бараг төгс давхцаж байгааг бид харж байна (3300 ба 3350 жил, ялангуяа тооцооллын алдааны хязгаарт). Энэ нь танилцуулсан гаплотип нь долоон гаплотипийн энэ салбарт бараг төгс нийцэж байгаа бөгөөд гаплотип, түүнээс хол зайд, мөн салбарын нийтлэг өвөг дээдсийн тооцоолсон насны хувьд тохирч байна гэсэн үг юм. Тиймээс бид ID00019 гаплотипийн салбарыг олсон. 39, 44, 45, 46, 51, 52 дугаартай холбогдох (ДНХ-ийн удам угсааны утгаараа) гаплотипууд аль дэд бүлэгт хамаарахыг харцгаая. Гаплотип 39 (Англи) нь танилцуулсан гаплотиптэй хамгийн ойр байдаг. Тэрээр 3100 жилийн настай P95 snip-тэй (энэ материалын эхэнд байгаа диаграммд шараар тодруулсан). Энэ нь хэлэлцэж буй салбарын нас буюу 3350±425 жилтэй бараг давхцаж байна. Дараа нь 51 ба 52 гаплотипууд (Шотланд, Швед) гарч ирдэг бөгөөд тэдгээр нь ID00019 шиг M223-аас илүү гүнзгий SNP-д зориулагдаагүй болно. Гэхдээ төсөл нь тэдгээрийг Z161, L801, CTS6433-аас ихгүй SNP гэж тодорхойлдог. Салбарын сүүлийн гурван гаплотип буюу 44, 45, 46 (Англи, Португал, Швейцарь) нь 2400, 3800, 3800 жилийн настай SNP Z79, CTS6433 болон CTS6433-тай. Тиймээс ID00019 салбар нь харьцангуй залуу бөгөөд SNP-ийн насыг тодорхой алдаагаар тодорхойлдог болохыг харгалзан түүн дээрх SNP-ууд нь насны хувьд тохирч байна. Дүгнэж хэлэхэд, ID00019 snips нь Тайлбарын эхэнд байгаа snip диаграммд шараар заасан зүсэлтүүдийн хүрээнд байх ёстой бөгөөд хамгийн их магадлалтай нь P95, CTS6433, Z79 снайпуудын дунд байх ёстой гэж дүгнэх хэрэгтэй. Москвагийн ДНХ-ийн удмын сангийн лаборатори эдгээр бүх SNP-ийг тодорхойлохоор төлөвлөж байна. Үзүүлсэн гаплотип хүртэлх түүхэн зам нь ойролцоогоор дараах байдалтай байна. Хаплогрупп I нь 43 мянган жилийн өмнө (IJ нэгдсэн гаплогруппээс) үүссэн бөгөөд 28 мянган жилийн өмнө I1 ба I2 гаплогруппуудад хуваагдаж, сүүлчийнхээс (мөн эхнийхээс) олон доод дэд бүлгүүд үүссэн. I2 хаплогруппын цаашдын түүхийг өмнөх тайлбаруудад тайлбарласан болно; Үүний үр дүнд Зүүн Европоос гаралтай I2a гаплогруппын ихэнх тээвэрлэгчид нь ойролцоогоор 2300 жилийн өмнө МЭӨ 1-р мянганы төгсгөлд нийтлэг өвөг дээдэс нь амьдарч байсан L147.2 дэд бүлгийн удам салбаруудад хамаардаг. ID00029 нь өөр нэг зэрэгцээ салбар болох M223-ын төлөөлөгч боловч нийтлэг өвөг дээдэс нь МЭӨ 2-р мянганы хоёрдугаар хагаст, ойролцоогоор 3300 жилийн өмнө амьдарч байжээ. Өргөдөл. Энд бид ID00029 салбараас гаплотип тээгчдээс олдсон SNP-ийн найрлагын талаарх дэлгэрэнгүй геномын мэдээллийг өгдөг. Энэ нь терминалын snips-ийг эс тооцвол бараг ижил байх ёстой. Гэхдээ дээр дурдсан хэд хэдэн тохиолдлуудын адил эдгээр нь адилхан байж магадгүй юм. M223 ба түүнээс доошхи SNP-уудыг тодруулсан. Иж бүрдэл дугаар 227059 (Англи, салбар дээрх гаплотип 39): CTS10057+, CTS10058+, CTS10100+, CTS10125+, CTS10362+, CTS109+, CTS11358+, CTS11441+, CTS1158+, CTS1123+, CTS116+, CTS115+ 2+, CTS1977+, CTS1996+, CTS2193+, CTS2392+, CTS2514+, CTS2536+, CTS3296+, CTS3326+, CTS3331+, CTS3431+, CTS3517+, CTS3536+, CTS3654+, CTS3662+, CTS3868+, CTS3996+, CTS4039+, CTS4088+, CTS429+, CTS44+, CTS431+, CTS4+, C34+ 8+, CTS444 3+, CTS4740+, CTS4848+, CTS4982+, CTS5286+, CTS5318+, CTS5457+, CTS5532+, CTS5650+, CTS5727+, CTS5908+, CTS6135+, CTS6136+, CTS616+, CTS6265+, CTS6331+, CTS6383+, CTS674+, CTS6800+, CTS6907+, CTS373+, CTS3727 +, CTS7682+, CTS7831 +, CTS7865+, CTS7922+, CTS7933+, CTS7934+, CTS8243+, CTS8333+, CTS8345+ , CTS8420+, CTS8449+, CTS88+, CTS8876+, CTS8901+, CTS8980+, CTS9183+, CTS9240+, CTS9264+, CTS9266+, CTS9482+, CTS9782+, CTS9782+, F102, F19+, F19+, F19+ , F1320+, F1329+, F14 50+, F1460+, F1704+, F1714+, F1753+, F1767+ , F2048+, F2075+, F2142+, F2155+, F2302+, F2345+, F2366+, F2402+, F2587+, F2688+, F2710+, F2794+, F2837+, F2794+, F2837+, F2315+, F263, F315+ 35+, F3368+, F3402+, F3556 +, F3692+, F4188+, F719+, F922+, L132+, L15+, L16+, L34+, L35+, L350+, L37+, L403+, L460+, L468+, L470+, L498+, L578+, L59+, L68+, L748+, L7+, L7+, L7+, L7+ , L800+, M139+, M168+, M17 0+, M223+, M235+, M294+, M42+, M89+, M94+, P123+, P124+, P126+, P127+, P130+, P135+, P136+, P138+, P14+, P141+, P145+, P14+, P15+, P118+, P15+, P118+ P160+, P166+, P187+, P214+ , P215+, P216+, P217+, P219+, P220+, P221+, P222+, P223+, P305+, P95+, PAGES00026+, PAGES00081+, PF1016+, PF1029+, PF101+, PF1029, P140, P4+ 061+, PF1092+, PF1097+, PF110+, PF1203+, PF1269+, PF1276+ , PF192+, PF210+, PF212+, PF223+, PF234+, PF258+, PF2591+, PF2593+, PF2599+, PF2608+, PF2611+, PF2615+, PF26+, PF26+, PF26+, PF26+, PF26+ F2745+, PF2747+, PF2748+, PF2749+, PF2770+, PF278+, PF292+, PF316+, PF325+ , PF342+, PF3515+, PF3517+, PF3518+, PF3534+, PF3560+, PF3561+, PF3562+, PF3573+, PF3574+, PF3578+, PF3586, P350F, PF3586, P350F 3595+, PF 3596+, PF3600+, PF3604+, PF3605+, PF3607+, PF3611+, PF3612+, PF3616+ , PF3618+, PF3621+, PF3623+, PF3625+, PF3626+, PF3634+, PF3639+, PF3640+, PF3641+, PF3642+, PF3644+, PF365+, PF365+, PF365, P365+, P365+ F3658+, PF3660+, PF 3664+, PF3666+, PF3667+, PF3669+, PF3671+, PF3672+, PF3675+ , PF3677+, PF3686+, PF3694+, PF3780+, PF3798+, PF3800+, PF3801+, PF3802+, PF3804+, PF3806+, PF3807+, PF3807+, PF3804, P38+, PF3801F, P3801F F3815+, PF3819+, PF3820+, PF 3822+, PF3836+, PF3837+, PF3849+, PF3876+, PF500+ , PF667+, PF719+, PF725+, PF779+, PF796+, PF803+, PF815+, PF821+, PF840+, PF844+, PF892+, PF937+, PF951+, PF937+, PF951+, PF68+, PF68+, V, PF17+ +, V205+, V221+, V241+, V250+, V52 +, V9+, YSC0000056+, YSC0000256+, YSC0000260+, YSC0000265+, YSC0000267+, YSC0000272+, YSC0000280+, YSC0000281+, YSC0000281+, YSC0000280+, YSC00000200+ Z161+, Z162+, Z163+, Z164+, Z165+, Z168+, Z170+, Z172+, Z174+, Z175+, Z176+, Z177+, Z178+, Z179+, Z181+, Z183+, Z184+, Z18, Z18+, Z17, Z18+ 0057+, CTS10058+, CTS10100+, CTS10125+ , CTS10834+, CTS11441+, CTS1977+, CTS2193+, CTS2392+, CTS2514+, CTS3296+, CTS3326+, CTS3517+, CTS3654+, CTS4039+, CTS444+, CTS47+, CTS448+, CTS438+ CTS4848+, CTS 4982+, CTS5286+, CTS5650+, CTS5727+, CTS5884+, CTS5908+, CTS6136+, CTS616+ , CTS623+, CTS6265+, CTS6331+, CTS674+, CTS6932+, CTS7329+, CTS7331+, CTS7682+, CTS7831+, CTS7865+, CTS7934+, CTS8333+, CTS845+, CTS8333+, CTS84+, CTS84+, 8+, CTS8876+, CTS890 1+, CTS9183+, CTS9240+, CTS9264+, CTS9266+, CTS9482+, CTS9782+ , F1209+, F3692+, F719+, L132+, L15+, L16+, L181+, L34+, L35+, L350+, L36+, L37+, L403+, L41+, L460+, L468+, L59+, L47+, L58+ 48+, L751+, L755+, L756+ , L758+, L772+, L800+, M168+, M170+, M213+, M223+, M235+, M258+, M294+, M299+, M42+, M438+, M89+, M94+, P123+, P12+, P12+, P12+, P12+, P12+, P12+, P12+ +, P130+, P133+, P134+ , P135+ , P136+, P138+, P139+, P14+, P140+, P141+, P143+, P145+, P146+, P148+, P149+, P151+, P157+, P158+, P159+, P161+, P160, P1+, P167, P1+ 12+, P214+, P216+, P21 7+ , P218+, P219+, P220+, P221+, P222+, P223+, P316+, P38+, P95+, PAGES00026+, PAGES00081+, PF2591+, PF2608+, PF2611+, PF2615+, PF2747+, PF2748+, PF2749+, PF2770+, PF3562+, PF6+4, P637F, P630F +, PF3641+, PF3644+, PF3651+, PF3652+, PF3657+, PF3658+, PF3660+, PF3664+, PF3666+, PF3667+, PF3669+, PF3671+, PF3672+, PF3675+, PF3677+, PF3780+, PF3800+, PF3801+, PF3802+, PF3804+, PF3809+, P381F, P381F, P3+1F 815+, PF 3819+, PF3820+, PF3822+, PF3836+, PF3837+, PF3849+, PF3876+, PF6464+, PF6469+, PF6470+, PF6477+, PF6479+, PF6520+, V241+, V250+, YSC0000207+, YSC0000227+, YSC0000256+, YSC00002702+, YSC00002701+, Z161+, Z163+, Z164+, Z165+, Z168+, Z170+, Z174+, Z175+, Z176+, Z178+, Z179+, Z184+, Z186+, Z188+, P37-, S2348-, S4442- Иж бүрдэл дугаар 173448 (Англи, гаплотип 44): Z77+, L800+, L801+, V218+, Z186+, Z78+, Z79+, L1198+, Z190+, Z185+, Z171+, Z166+, CTS10057+, CTS10058+, CTS10100+, CTS10125+, CTS10834+, CTS11441+, CTS2134+, CTS2193+, CTS2199+, C33+, CTS3+, CTS29 26+, CTS3517+, CTS3654+, CTS4039+, CTS4088+, CTS429+, CTS4348+, CTS4437+, CTS4848+ , CTS4982+, CTS5286+, CTS5650+, CTS5727+, CTS5884+, CTS5908+, CTS6136+, CTS616+, CTS623+, CTS6265+, CTS6331+, CTS6433+, CTS674+, CTS6932+, CTS7329+, CTS7331+, CTS7682+, CTS7831+, CTS7865+, CTS7934+, CTS8333+, CTS8345+, CTS8420+, CTS8449+, CTS8819, CTS88+, CTS88+, CTS88+, 3+, CTS9240+, CTS9264+, CTS9266+, CTS9482+, CTS9782+, F1209+, F3692+, F719+ , L104+, L132+, L15+, L16+, L181+, L34+, L35+, L350+, L36+, L37+, L403+, L41+, L460+, L468+, L498+, L578+, L59, L57, L57, L57, L57 +, L758+, L772+, L800+ , M168+, M170+, M213+, M223+, M235+, M258+, M294+, M299+, M42+, M438+, M89+, M94+, P108+, P123+, P124+, P125+, P126+, P127+, P129+, P130+, P13+, P13+, P13+, P13+, P13+, P13+ , P139+, P14+, P140+, P141+ , P143+, P145+, P146+, P148+, P149+, P151+, P157+, P158+, P159+, P160+, P161+, P163+, P166+, P187+, P212+, P187+, P212+, P214, P2+, P218, P2+ 20+, P221+, P222+, P223+ , P305+ , P316+, PAGES00026+, PAGES00081+, PF2591+, PF2608+, PF2611+, PF2615+, PF2624+, PF2747+, PF2748+, PF2747+, PF2749, P3+, P3749F F3626+, PF3639+, PF3640+, PF3641+, PF3644+, PF3651+, PF3652+, PF3657+, PF3658+, PF3660+ , PF3664+, PF3666+, PF3667+, PF3669+, PF3671+, PF3672+, PF3675+, PF3677+, PF3780+, PF3800+, PF3801+, PF3800+, PF3801+, PF3802, PF380+, PF3802, P380+ F3812+, PF 3814+, PF3815+, PF3819+, PF3820+, PF3822+, PF3836+, PF3837+, PF3849+ , PF3876+, PF6464+, PF6469+, PF6470+, PF6477+, PF6479+, PF6520+, PK1+, SRY10831+, V221+, V241+, V250+, YSC002, YSC02000+, YSC002, YSC02000 YSC0000272+, Z163+, Z164+, Z165+, Z168+, Z170+, Z174+, Z175+, Z178+, Z179+ , Z184+, Z185+, Z188+, Z190+, Z79+, L707+, M223+, M253-, M26-, M284-, M379-, P78-, P95-, S2348-, S4442- Иж бүрдэл дугаар 18187 (Португаль, гаплотип 45): CTS674+, CTS6800+, CTS6907+, CTS6932+, CTS7329+, CTS7331+, CTS7502+, CTS7682+, CTS7831+, CTS789+, CTS379+, CTS379+ TS 8243+, CTS8333+, CTS8345+, CTS8420+, CTS8449+, CTS88+, CTS8876+, CTS8901+, CTS8980+, CTS9183+, CTS9240+, CTS9264+, CTS9266+, CTS9482+, CTS9782+, CTS9828+, F1046+, F1209+, F1302+, F1209+, F1302+, F1304+, F1104+ 04+, F171 4+, F1753+, F1767+, F2048+, F2075+, F2142+, F2155+, F2302+, F2345+, F2366+, F2402+, F2587+, F2688+, F2710+, F2794+, F2837+, F2985+, F2993+, F3111+, F3136+, F336+, F336+, F336+, F336+, F365, F365 +, F4188+, F719+, F922+, L132+, L15+, L16+, L34+, L35+, L350+, L37+, L403+, L41+, L460+, L468+, L470+, L498+, L578+, L59+, L68+, L748+, L751+, L755+, L756+, L755+, L756+, L7728, M17+, M17+, L17+, M17+, L17+, L460+, L468+, L470+, L709 0+, M223+, M235+, M294+, M42+, M89+, M94+, P123+, P124+, P126+, P127+, P130+, P135+, P136+, P138+, P14+, P141+, P145+, P14+, P15+, P118+, P15+, P118+ P160+, P166+, P187+, P214+ , P215+, P216+, P217+, P219+, P220+, P221+, P222+, P223+, PF1016+, PF1029+, PF1031+, PF1040+, PF1046+, PF1046+, PF1061F, P1710F, PF+ 203+, PF1269+, PF1276+, PF192+, PF210+, PF212+, PF223+ , PF234+, PF258+, PF2591+, PF2593+, PF2599+, PF2608+, PF2611+, PF2615+, PF2624+, PF263+, PF2643+, PF272+5, PF24+, PF24, P7, P7 9+, PF277 0+, PF278+, PF292+, PF316+, PF325+, PF342+, PF3515+, PF3517+ , PF3518+, PF3534+, PF3560+, PF3561+, PF3562+, PF3573+, PF3574+, PF3578+, PF3586+, PF3588+, PF3590+, PF359+, PF359+, P3F604, P3F+, P3F605 F3604+, PF3605+, PF 3607+, PF3611+, PF3612+, PF3616+, PF3618+, PF3621+, PF3623+ , PF3625+, PF3626+, PF3634+, PF3639+, PF3640+, PF3641+, PF3642+, PF3644+, PF3651+, PF3652+, PF3654+, PF3654+, PF365, P36+, P365, P36+, P3F64F F3666+, PF3667+, PF3669+, PF 3671+, PF3672+, PF3675+, PF3677+, PF3686+, PF3694+ , PF3780+, PF3798+, PF3800+, PF3801+, PF3802+, PF3804+, PF3806+, PF3807+, PF3809+, PF3811+, PF3812+, PF3814, P38+, PF3814, P381F F3822+, PF3836+, PF3837+, PF3849+, PF 3876+, PF500+, PF667+, PF719+, PF725+ , PF779+, PF796+, PF803+, PF815+, PF821+, PF840+, PF844+, PF892+, PF937+, PF951+, PF954+, PF970+, V186+, V25, V180+, V25, V180+ , YSC0000256+, YSC0000260+, YSC0000265+, YSC 0000267+, YSC0000272+, YSC0000280+ , YSC0000281+, YSC0000298+, YSC0000300+, Z161+, Z162+, Z163+, Z164+, Z165+, M170+, M223+, M258+, P38+, L801+, Z186+, Z76+, L1316+, CTS10057+, CTS10058+, CTS10058+, CTS1010+, CTS1010+, CTS1010+, CTS1019+ , CTS11358+, CTS11441+, CTS11575+, CTS11726+, CTS125+, CTS12632+, CTS1996+ , CTS2193+, CTS2392+, CTS2514+, CTS2536+, CTS3296+, CTS3326+, CTS3331+, CTS3431+, CTS3517+, CTS3536+, CTS3654+, CTS3662+, CTS3689+, CTS3689, CTS34+, CTS3689+ 4088+, CTS429+, CTS4314+, CTS4348+, CTS4364+, CTS4368+, CTS4443+, CTS4740+, CTS4848+ , CTS4982+, CTS5286+, CTS5318+, CTS5457+, CTS5532+, CTS5650+, CTS5727+, CTS5908+, CTS6135+, CTS6136+, CTS616+, CTS6265+, CTS6263+, CTS6263+, CTS6433+, F3692+, F719+, L104+, L132+, L15+, L16+, L181+, L34+, L35+, L350+, L36+, L37+, L403+, L41+, L460+, L468+, L498+, L15+, L57, L6+ +, L756+, L758+, L772+, L800+, M168+, M170+, M213+, M223+, M235+, M258+, M294+, M299+, M42+, M438+, M89+, M94+, P123+, P124+, P12+, P12+, P116+, P125 P133+ , P134+, P135+, P136+, P138+ , P139+, P14+, P140+, P141+, P145+, P146+, P148+, P149+, P151+, P157+, P158+, P159+, P160+, P161+, P163+, P16+, P12+, P16+, P12+, P16+, P12 +, P21 7+, P218+, P219+, P220+, P221+, P222+, P223+, P316+, PAGES00026+, PAGES00081+, PF2591+, PF2608+, PF2611+, PF2615+, PF2747+, PF2747+, PF2747, P4704F 3833+, Z2198 +, CTS8444+, CTS6497+, CTS10057+, CTS10058+, CTS10100+, CTS10125+, CTS10834+ , CTS11441+, CTS2134+, CTS2193+, CTS2392+, CTS2514+, CTS3296+, CTS3326+, CTS3517+, CTS3654+, CTS4039+, CTS4088+, CTS424+, CTS424+, CTS424+ TS4982+, CTS5286+, CTS5 650+, CTS5727+, CTS5884+, CTS5908+, CTS6136+, CTS616+, CTS623+ , CTS6265+, CTS6331+, CTS674+, CTS6932+, CTS7329+, CTS7331+, CTS7682+, CTS7831+, CTS7934+, CTS8333+, CTS8345+, CTS8420+, CTS8420+, CTS8420+, CTS888+ 01+, CTS9183+, CTS9240+, CTS92 64+, CTS9266+, CTS9482+, CTS9782+, PF3562+, PF3574+ , PF3626+, PF3639+, PF3640+, PF3641+, PF3644+, PF3651+, PF3652+, PF3657+, PF3658+, PF3660+, PF3664+, PF366+, PF366, P366+, P3F66, P3F66+ F3672+, PF3675+, PF3677+, PF3780+, PF 3800+, PF3801+, PF3802+, PF3804+, PF3809+ , PF3811+, PF3812+, PF3814+, PF3815+, PF3819+, PF3820+, PF3822+, PF3836+, PF3837+, PF3849+, PF3876+, Z172, Z17+, Z17, Z17+, Z17+, Z17, Z17+ 8+, Z179+, P19+, Z181+, Z183+, Z184+, Z186+, V22 1+ , V241+, V250+, YSC0000207+, YSC0000227+, YSC0000256+, YSC0000272+, Z163+, Z164+, Z165+, Z168+, Z170+, Z174+, Z6+, Z6+, Z18F 469+, PF6470+, PF6477+, PF6479+, PF6520+, Z168+, Z170+, Z188+, Z7 7+, Z188+, F1209+, F3406-, M161-, Z190 -, Z63-, Z79-, L1317-, Z171-, S2348-, S4442-, Z185-, Z187-, L1201-, P30-1, P30, -, PF4225-, DF5-, PF3292-, CTS661-, L1198-, Z190-, Z2059-, Z2084-, Z2074-, L1272-, ZS6-, ZS7-, ZS11-, ZS14-, ZS15-, ZS16-, ZS18-, ZS26-, L1290-, Z78-, Z79-, M284-, M379-, P78-, P95-, L380-, M26-, M307-, M72-, M227-, M253-, M21-, Z166- Иж бүрдэл дугаар B3411 (Швейцарь, гаплотип 46): L801+, CTS6433+, M223+, L1201-, Z190-, Z78-, CTS5332- Иж бүрдэл дугаар 23157 (Шотланд, гаплотип 51): P19+, M170+, M258+, M223+, P19+, P38+, M284-, M379-, P78-, P95-, P30-, P37-, M227-, M253-, M26-, M307-, M72-, M21-, M161- Иж бүрдэл дугаар 174472 (Швед, гаплотип 52): M284-, L126-, M379-, P78-, P95- Haplogroup I2c KLIN ID00005 FTDNA-ийн илүү гүн дэд бүлэг болох I2c-L596 дэд бүлэг нь тогтоогдоогүй байна. 14 24 15 10 12 13 11 13 12 12 11 28 – 18 8 8 11 11 24 15 21 31 11 11 15 15 – 10 10 19 21 15 13 17 17 34 34 14 10 12 8 16 16 8 13 10 8 10 9 12 21 21 16 11 12 13 13 8 14 25 21 12 13 11 13 11 12 12 11 Дэд бүлэг нь I2c-L596-аас илүү гүн тодорхойлогдоогүй тул I гаплогруппын бодит тодорхойлсон дэд бүлгүүдийн гинж нэлээд богино байна: I-M170 > I2-M438 > I2c-L596. FTDNA-д таних нь зөв хийгдсэн бөгөөд энэ нь 67 маркер гаплотипийн мод дээрх танилцуулсан гаплотипийн байрлалаас харагдаж байна. Гэхдээ хараахан тогтоогдоогүй байгаа дараах буурах дэд бүлгүүд боломжтой.
Одоо 9 гаплотипийн салааны тухай (Гүржээс гурав нь [427, 451, 452 дугаартай], Беларусь хоёр, Литва, Австри, Туркээс тус бүр нэг). Энэ салбарын үндсэн гаплотип (танилцуулсан гаплотипээс мутацийн ялгааг тэмдэглэв) 14 24 15 10 12 13 11 13 11 13 11 29 – 18 8 8 11 11 24 15 20 32 11 14 15 16 – 11 10 19 21 15 13 17 19 34 36 12 10 12 8 16 16 8 12 10 8 10 9 12 21 21 16 11 12 13 14 8 13 25 20 12 13 11 13 11 12 12 11 Салбарын бүх 9 гаплотип нь энэхүү үндсэн гаплотипээс 123 мутацитай бөгөөд энэ нь салааны нийтлэг өвөг 123/9/0.12 = 114 → 129 ердийн үеийн өмнө буюу 3225 ± 430 жилийн өмнө амьдарч байсан гэсэн үг юм. Үзүүлсэн гаплотип нь үндсэн нэгээс 17 мутациар ялгаатай (олон тооны мутацыг тусгай дүрмийн дагуу тооцдог) бөгөөд энэ нь тооцооллын алдааны дотор нийтлэг өвөг дээдсээс ойролцоогоор ижил цагийг өгдөг. Энэ нь ID00005 нь МЭӨ 2-р мянганы төгсгөлд амьдарч байсан I2c хаплогруппын салбарын нийтлэг өвөг дээдсийн шууд удам болохыг харуулж байна.
L596 дэд бүлэг ба түүний доод дэд бүлгүүдийн түүх хичнээн төвөгтэй, эмгэнэлтэй байсныг ойлгохын тулд L596 дэд бүлэг нь өөрөө ойролцоогоор 21,300 жилийн өмнө үүссэнийг (Y хромосом дахь SNP-ийн шатнаас харж болно) тэмдэглэж байна. Европын түүхэнд, дараа нь L596-ийн амьд үлдсэн цөөхөн тээвэрлэгчдээс дахин төрсөн. Үүний үр дүнд ID00005 дэд салбаруудын өвөг дээдэс ойролцоогоор 3225 жилийн өмнө амьдарч байсан бол дээрх диаграмм дахь хөрш салбар (долоон гаплотип) - ойролцоогоор 2475 жилийн өмнө, дээд салбар (есөн гаплотип) - 2800 жилийн өмнө, мөн бүх гурван салбарууд амьдарч байжээ. хамтдаа - 5950 ± 645 жилийн өмнө. Мөн геномын шинжилгээ нь доод дэд бүлэг I2c1 нь 15,500 жилийн өмнө үүссэн болохыг харуулж байна; доорх дараагийн дэд бүлэг, I2c1a-L1251 - 10,100 жилийн өмнө, үлдсэн тухай мэдээлэл хараахан алга байна. Өнөөг хүртэл L596-ийн гурван салбар нээгдсэнийг бид харж байгаа бөгөөд тус бүр нь 3200-2500 жилийн өмнө үүссэн.
Ингээд өнөөдрийн мэдлэгийнхээ төлөв байдлыг нэгтгэн дүгнэе. Хаплогрупп I нь маш эртний бөгөөд 40 мянга гаруй жилийн өмнө үүссэн бөгөөд одоо хүртэл тодорхойгүй байгаа боловч Европт байх магадлалтай. 30 мянга орчим жилийн өмнө энэ нь I1, I2 дэд бүлэгт хуваагдаж, сүүлийнх нь ойролцоогоор 21,300 жилийн өмнө I2a, I2b, I2c-L596 дэд бүлэгт хуваагджээ. Хаплогрупп I ба I2 нь 7 мянган жилийн өмнөх Төв Европ, Шведэд эртний ясны олдворуудаас олдсон. Хэсэг хугацааны дараа тэнд гурван салааны нийтлэг өвөг дээдэс I2c-L596 амьдарч байсан бөгөөд түүний үр удам нь одоогоор Британийн арлуудаас Турк хүртэл амьдардаг. Гэвч 4500 орчим жилийн өмнө хаплогрупп I тээгчдийн амьдрал Европ дахь бусад хүмүүсийн нэгэн адил эрс тасалдаж, бараг бүх гаплогруппууд (R1b-ээс бусад) Европоос алга болж, эсвэл Европын захын бүс нутгууд руу буцаж очсон. Баруун Ази, Кавказ. Эдгээр гаплогруппууд нь бие даасан төлөөлөгчдийн хувьд амьд үлдэх урт, зовлонтой үеийг туулж, амьд үлдсэн хэсэг нь үржиж, шинэ салбаруудын нийтлэг өвөг дээдсийг бүрдүүлдэг. Тэдний дунд 3200-2500 жилийн өмнө, өөрөөр хэлбэл МЭӨ 2-р зууны төгсгөл - 1-р мянганы дунд үеэс "өсч" эхэлсэн I2c-L596 дэд бүлгийн тээвэрлэгчид байв. Бараг нэгэн зэрэг I2a хаплогрупп сэргэж эхэлсэн бөгөөд нийтлэг өвөг дээдэс нь Зүүн Европт 2300 жилийн өмнө, 5000 орчим жилийн өмнө Британийн арлуудад амьдарч байжээ. Хэсэг хугацааны өмнө I1 хаплогрупп сэргэж эхэлсэн бөгөөд ойролцоогоор 3700 жилийн өмнө нийтлэг өвөг дээдэс нь амьд үлджээ. Үүнтэй холбогдуулан I2c-L596 дэд бүлэг нь 6000 орчим жилийн өмнө энэ гаплогруппын өнөөгийн тээвэрлэгчидтэй холбоотой хамгийн эртний үндэстэй юм. L596-ийн доод дэд бүлгүүд нь гаплотипийн харгалзах салбаруудад хуваагдсан нь таатай байна (дээрх диаграмыг үз). Энэ нь эдгээр доод дэд бүлгүүдийг Москвагийн ДНХ-ийн удмын бичгийн лабораторид хангалттай тооны (жишээлбэл, тавин) хүнээр тодорхойлох үед бид гаплотипуудыг дэд бүлэгт илүү нарийвчилсан хуваарилалтыг хийж, илүү нарийвчлан ойлгох боломжтой болно гэсэн үг юм. дэд бүлгүүдийн зонхилох газарзүй. Хувийн mtDNA тайлбарын тест Эмэгтэй гаплогруппуудад зориулсан хувийн тайлбарын хүрээг өргөжүүлэх нь мэдээж хэрэг бөгөөд эмэгтэйчүүд, эрэгтэйчүүдийн mDNA-г шинжлэхийн тулд шинжилж болно. Үнэн хэрэгтээ эр хүн эхийнхээ цусан дахь mtDNA-г хүлээн авдаг боловч mtDNA-г цааш дамжуулдаггүй, учир нь эр бэлгийн эсийн митохондри нь бордсоны дараа (эсвэл түүний явцад) устдаг. Эр бэлгийн эс нь арав орчим митохондри агуулдаг, учир нь сүүлний эргэлт нь эрчим хүч шаарддаг бөгөөд үүнийг митохондри хангадаг. Эр бэлгийн эс зорилгодоо хүрэхэд митохондриа хаядаг. Та эрчүүдийг таньж байна уу? Маш ховор тохиолдолд митохондри нь хальтирсан хэвээр байгаа боловч энэ нь жирэмсэн эхийн биетэй холбоотой гадаад митохондри бөгөөд энэ нь дүрмээр бол сайнаар төгсдөггүй. Мөн энэ нь ихэвчлэн цочмог эмгэгээр төгсдөг. Товчхондоо, mtDNA нь эхээс охидод үе дамжсан, хэдэн арван, хэдэн зуун мянган жил, сая сая жил гэх мэт бүхэл бүтэн гинжин хэлхээний дагуу дамждаг. Мөн хөвгүүдийн хувьд - зөвхөн нэг үеийнхэн. Хөвгүүд нь Y хромосомыг хөвгүүддээ дамжуулдаг, mtDNA-г хэнд ч дамжуулдаггүй - хөвгүүд ч, охид ч биш, харин бусад 22 хромосомыг рекомбинант болгон хүн бүрт дамжуулдаг. Тиймээс би давтан хэлье, mtDNA шинжилгээг эмэгтэй, эрэгтэй хүмүүсийн ДНХ дээр хийж болно. Аль ч тохиолдолд үр дүн нь ижил байна - эхийн mtDNA болон өмнөх бүх mtDNA гинжийг бүх хуримтлагдсан мутацитай тодорхойлох. Энэ нь юу өгдөг талаар олон хэвлэлд, ялангуяа бидний хувьд хамгийн тохиромжтой хэлбэрээр - "Интернет. Судлаачийн тэмдэглэл" (2010), 368-379-р хуудас, бага зэрэг өргөжүүлсэн хэлбэрээр - "Славуудын үүсэл" (2013), 265-284-р хуудас. mtDNA-г олон нийтэд тайлбарлах кампанит ажлыг эхлүүлэхийн өмнө бид нэг хамтрагчийн хийсэн туршилтын хувилбарыг санал болгож байна. Энэ нь туршилтын хувилбар тул тайлбар зохиогчийн нэрийг одоогоор нуусан байна. Энэ үе шатанд бүтээгдэхүүнийг хамгийн багадаа хүлээн зөвшөөрч болохуйц, илүү сайн, сайн эсвэл бүр маш сайн гэж үзэх нь чухал юм. Тиймээс энэ нь боломжит үйлчлүүлэгчид, түүнчлэн бүх уншигчдад зориулагдсан юм. Тиймээс туршилтын сонголтын хувьд дараахь зүйлийг анхаарч үзээрэй.
Энэхүү өгөгдлүүд нь уг угсаатны түүхийг палеолит (чулуун зэвсгийн үе) -ээс МЭӨ хэдэн мянган жилийн тэнгэрийн хаяа хүртэлх гүнд судлах боломжийг бидэнд олгодог. Энэ нь нэгдүгээрт, эмэгтэй митохондрийн ДНХ нь маш их хувьсах бүс нутгуудаас хамаагүй богино урттай байдаг тул мэдээлэл багатай байдагтай холбоотой юм. Хоёрдугаарт, танилцуулсан өгөгдөл нь хүртээмжтэй нарийн ширийн зүйлийг агуулаагүй болно - жишээлбэл, доор харуулсны дагуу танилцуулсан мутацийн зурагт хамаарах H1c дэд бүлэг нь 20 гаруй доод дэд бүлгийг агуулдаг. Тиймээс уг удам угсааны түүхийг зөвхөн Y хромосомын талаарх өгөгдлийн тайлбартай харьцуулахад илүү их ойролцоо байдлаар сэргээж болно. HVR1 ба HVR2-ийн RSRS-тай ялгааг шинжлэх нь танилцуулсан гаплотип нь митохондрийн гаплогрупп H1c-д хамаардаг болохыг харуулж байна. Дээр дурдсан шалтгааны улмаас өгсөн дээж нь бусад олон mtDNA-ийн өгөгдөлтэй таарч байна: familytreedna.com мэдээллийн санд HVR1 ба HVR2-ийн ижил RSRS утгатай гаплотипуудын тээвэрлэгчид байдаг бөгөөд тэдгээрийн дийлэнх нь хойд болон зүүн хэсэгт хамаардаг. Жишээлбэл, Европ, mtDNA-д зориулсан Anela Sarocka (Литва). Хаплотипийн түүх. Эмэгтэй гаплогрупп H нь 30 орчим мянган жилийн өмнө Баруун Азид үүссэн, 20-25 мянган жилийн өмнө Европт орж ирсэн бөгөөд тивийн баруун өмнөд хэсэгт Франц-Кантабрийн бүс нутагт тархсан гэж үздэг бөгөөд энэ нь Мадлений соёлтой нийцдэг. . 20-13 мянган жилийн өмнөх хамгийн сүүлчийн мөстлөгийн үеэр Хойд ба Төв Европын палеолитын ихэнх суурингууд устаж үгүй болсон тул H гаплогруппын төлөөлөгчид зөвхөн Испанийн хойд хэсэгт л амьд үлджээ (тиймээс одоогийн байдлаар энэ гаплогрупп өндөр давтамжтай, 50% -иас илүү нь энэ бүс нутагт Хятад-Кавказын харь гарагийн оршин суугчдыг уусгах явцад үүссэн Баскуудын дунд олддог). Энэ (чулуужсан) гаплогруппыг Ла Пасьега, Ла Чора (мадлен) агуйд илрүүлсэн. Хожуу тал дээр мезо-неолитын үеэс эхлэн Хойд Европ, Оросын тэгш тал дээр хаплогрупп эрт байгааг тэмдэглэх нь зүйтэй - Сертея (Смоленск муж), Сертея VIII давхаргад H гаплотипийн тээвэрлэгч олдсон. эр гаплотип R1a1-тэй хослуулан (МЭӨ IV мянган). Энэхүү археологийн соёлыг Юүлүүр аяганы соёлын алс хязгаар гэж үзэж болно. Түүнчлэн МЭӨ 5-4-р мянганы оршуулгын газраас ижил төстэй гаплотипууд олдсон. Донецк мужид болон Южный Олений Островын оршуулгын газарт (түүн дээр R1a1* эрэгтэй гаплогрупп бас тогтоогдсон). Дэд бүлэг H1c. T477C үзүүлэлт нь гаплотип нь H1c дэд бүлэгт хамаарах эсэхийг тодорхойлох боломжийг олгодог. H1, H3 дэд ангиуд, түүнчлэн эгч хаплогрупп V-ийн тархалт нь 13 мянган жилийн өмнөх сүүлчийн мөстлөгийн дараа Франц-Кантабрийн бүс нутагт Европ доторх тэлэлттэй холбоотой гэж таамаглаж байна. Хаплогрупп H1 нь Баруун Европын митохондрийн ДНХ-ийн нэлээд хувийг эзэлдэг бөгөөд Испанийн Баскууд (28%), Португальчууд (26%), Андалузын оршин суугчид (24%), Кантабриа дахь Пасиего угсаатны зүйн бүлгүүд голлон хуваалцдаг. (24%). Хойд Африкт Ливийн туарегуудын дунд H1 гаплогруппын эзлэх хувь ер бусын өндөр (61%) байгаа нь Испанитай зэргэлдээх бүс нутгийг мөн харуулж байна. Мөн mtDNA гаплогрупп H1 нь Иберийн хойг, Хойд Африк, Сардинийн бусад оршин суугчдын дунд түгээмэл байдаг. Түүгээр ч зогсохгүй Франц, Британийн арлууд, Альпийн нуруу, Зүүн Европын олон бүс нутгийн хүн амын 10 гаруй хувийг тээвэрлэгчид, Европын бусад газруудад дор хаяж 5 хувийг эзэлдэг. H1c гаплогруппын нас 9400 орчим жил байна. Үүнтэй холбогдуулан түүний Хойд болон Зүүн Европ руу нүүсэн цаг, огнооны тухай асуулт гарч ирнэ. Энэ дэд бүлгийг МЭӨ 3500-2500 оны үед Скандинавын Юүлүүр аяга соёлын тариаланчдын дунд нээсэн. (Gokhem2). Үүнтэй төстэй гаплотип нь Скандинав болон Хойд Европын өмнөх соёлд олдсонгүй. Мезо-неолитийн үед Европын хойд ба зүүн хэсэгт орших энэхүү дэд давхарга нь Олений арлаас Украйны өмнөд хэсэг хүртэл өргөн зурваст гарч ирснийг харгалзан үзвэл энэ нь өмнөд хэсгээс шилжин суурьших тухай асуудлыг хөндөх боломжийг бидэнд олгож байна. Франц-Испанийн бүс нутгийг өмнөх эрин үед энэхүү гаплогруппын тархалтын төвүүдийн нэг гэж үзэж болох тул энэхүү гаплотипийн харагдах байдлын талаархи логик таамаглалыг Хойд ба Зүүн Европ руу далайгаар нэвтрэх хувилбар гэж үзэж болно. Зүрхний керамик соёлын тээвэрлэгчдийн Газар дундын тэнгисийн сав газар (Д. Л. Гаскевичийн Хойд Понтийн импрессогийн талаархи таамаглал). Энэ нь Баруун Европоос гадуурх H1 тээвэрлэгчдийн дүр төрхийг Кардиак керамик цогцолборын шилжилт хөдөлгөөнтэй (эсвэл бусад соёл, археологийн холболттой) холбох боломжийг олгодог бөгөөд тэдгээрийн төлөөлөгчид арьсан завиар эрэг дагуу хөдөлж чаддаг. Гаскевич МЭӨ 6-р мянган жил хүртэл оршин тогтнож байсан Баруун Европ, Газар дундын тэнгисээс энэ дэд давхарга байдаг мегалит соёл, зүрхний керамик эдлэлийг бүтээгчдийн талаар ч дурджээ. Буг-Днестрийн соёлоор дамжин Крым болон өмнөд Украин руу Босфорын Истмус.
Шинжилгээний үр дүнд танилцуулсан гаплотип нь олон судлаачид Индо-Европ гэж үздэг Юүлүүр аяганы соёлын төлөөлөгчийн Гохем2-ээс олдсон дэд бүлэгтэй тохирч байгааг бид хэлж чадна. Эхийн H гаплогрупп нь R1a1 (Өмнөд Олений Остров, Сертея) эр гаплотипүүд байсан газраас олдсон бөгөөд энэ нь Оросын тал нутгийн уугуул, эрэгтэй R1a1-ийн нэгэн төрлийн "хамтрагч" гэж үзэх боломжийг бидэнд олгодог. Илүү урт холболтууд нь H1c-ийн үндэсийг Газар дундын тэнгис рүү чиглүүлдэг. Үйлчлүүлэгчийн удам угсааг нарийвчлан судлахын тулд илүү нарийвчилсан бичвэр хийхийг зөвлөж байна, учир нь H1c хаплогрупп нь 20 гаруй өөр дэд бүлгүүдийг агуулдаг тул бид танд сануулж байна. Ийм шивэх ажлыг Москвагийн ДНХ-ийн угийн бичгийн лаборатори удахгүй хийх болно. Haplogroup I2 нь 15 мянга гаруй жилийн өмнө зүүн өмнөд Европт үүссэн. Түүний хамгийн их концентраци нь Босни-Герцеговина, Далматийн өмнөд хэсэгт (Хорват) байдаг бөгөөд бүх Өмнөд Славян хүн амын нэлээд хувийг эзэлдэг.
Эдгээр бүсүүд дэх I2a-ийн өндөр нягтрал нь орчин үеийн Хорватын Адриатын бүс нутаг болон Динарийн Альпийн нуруу нь сүүлийн мөстлөгийн үед I2a-г тээгч палеопопуляцийн байгалийн хоргодох газар байсныг харуулж байна. I2a хаплогруппын төлөөлөгчид Балканы хойгийн хамгийн эртний соёлын гарал үүсэлтэй байсан, тухайлбал: Чел Кладове, Лепенски Вир, Гура Бациулуи, Неа Никомедиа. Энэ овгийн үндсэн дээр Иллири ба Фракийн овог аймгууд хөгжсөн бололтой. I2a салбар нь Сардинид анх суурьшсан салбаруудын нэг байв. Haplogroup салбарууд:I2a1Хаплогрупп I2a1(M26) Сардини дахь нийт эрэгтэй удмын 40 орчим хувийг эзэлдэг. Нэмж дурдахад энэ нь Баскийн улс болон Иберийн хойгт бага, дунд зэргийн давтамжтай байдаг бөгөөд Македон улсад амьдардаг албаничуудын түүвэрт 1.6% (1/64), түүвэрт 1.2% (3/257) тус тус эзэлж байна. Чехов M26 дэд бүлгийн YSTR өөрчлөлтийн нас нь 8.0 ± 4.0 мянган жил байна (Rootsi 2004). салбар гэж таамаглаж байна I2a-аас ялгарч байв I2 11,000 орчим жилийн өмнө Балканы хойгт, мөн салбар I2a1-аас ялгарч байв I2a 8000 орчим жилийн өмнө Сардинид эсвэл Европын эрэг орчмын хаа нэгтээ. I2a2 Хаплогруппын тархалт I2a2 I2a2(snp M423) нь зүүн өмнөд Европын хүн амын дунд түгээмэл байдаг. Хамгийн их нягтрал нь Далмати (Хорват), Босни-Герцеговина (>50%) байдаг. Динарийн өндөрлөг дээрх энэ хэсэгт энэ салбар үүссэн гэж таамаглаж байна I2a 7500 орчим жилийн өмнө. Хамгийн түгээмэл гаплогрупп I2a2Славян ард түмний дунд (ялангуяа Өмнөд Славян), мөн Румын, Молдав, Унгар, Литва, Албани, Грекийн өмнөд, Италийн зүүн хойд хэсэг, Баруун Анатоли, Хойд Кавказын оршин суугчдын дунд олддог. Хаплогруппын хамгийн өндөр давтамж, олон янз байдал I2a2Баруун Балканы хүн амын дунд орчин үеийн Хорватын Адриатын бүс нутаг сүүлчийн мөстлөгийн үед энэ гаплогруппын тээвэрлэгчдийн хоргодох газар байсан гэж үздэг. snp P37.2-ийн салбар нь 10.7 ± 4.8 мянган жилийн өмнө үүссэн (Rootsi 2004). snp P37.2 салбарын нас нь 8.0 ± 4.0 мянган жил (Rootsi 2004), snp M423-ийн хувьд 8.8 ± 3.6 мянган жил (Underhill 2007). Перисик энэ салбарын тархалт нь "Голоцений шилжилтээс өмнө биш, Эрт Неолитийн үеэс ч хожуу биш" (Pericic 2005) гэж үздэг. Төвлөрлийн хоёр дахь төв I2a2Одоогийн Молдав улсад байрладаг бөгөөд Трипиллийн соёлын хилтэй ойролцоогоор давхцаж байгаа тул энэ дэд бүлэг нь энэ соёлын онцлог шинж чанартай байсан бөгөөд Индо-Европчууд ирснээр тэд бусад салбаруудын адил дарангуйлалд өртөөгүй бөгөөд тэдний удмын санд нэгдсэн. гаплогрупп Iтөв болон баруун Европт. I2b1Дэд бүлгийн хуваарилалт I2b1гаплогруппын тархалттай давхцаж байна I1, Фенноскандиаг эс тооцвол энэ нь палеолитын зэвсгийн дор хаяж нэг хоргодох байранд байгааг илтгэж байгаа бөгөөд энд хаплогрупп бас байрладаг байв. I1. Байхгүй I2b1Фенноскандиад хаплогрупп байдагтай холбоотой байж болох юм I2b1Хаплогрупп үүссэнээс хойш энэ бүс нутгийн хамгийн эртний суурин газруудад "үүсгэн байгуулагчдын нөлөө" болон ховор тохиолдлын улмаас генетикийн шилжилт нөлөөлсөн. I2b1Энэ нь Доод Саксонийн гаднах заасан газар нутгийн нийт хүн амын Y-хромосомын олон янз байдлын 10% -иас бага хувийг бүрдүүлдэг. Гаплогруппуудын тархалт I1Тэгээд I2b1герман хэлээр ярьдаг хүмүүсийн тархалтын түүхэн хил хязгаартай харьцангуй сайн уялдаж байна. Хаплогрупп I2b1Зөвхөн Герман, Нидерланд, Бельги, Дани, Англи (Уэльс, Корнуоллаас бусад), Шотланд, Шведийн өмнөд хэсэг, Норвеги, Норманди, Мэн, Анжу мужуудад л хүн амын 4% -иас илүү байдаг. болон Францын баруун хойд хэсэгт Перче, зүүн өмнөд Франц дахь Прованс, Италийн түүхэн бүс нутагт - Тоскана, Умбрия, Латиум; түүнчлэн Молдав, Рязань муж, Мордовид. Орчин үеийн Франц, Англи, Итали, түүнчлэн зүүн Европт I1 ба I2b1 байгаа нь Кельт, Германы тэлэлттэй аль хэдийн холбоотой байсан бөгөөд Индо-Европоос өмнөх үед эдгээр гаплогруппууд зөвхөн хойд хэсэгт төвлөрч байсан бололтой. Европ. Хаплогруппын салбаруудын нэг I2b1, тухайлбал I2b1a(snp M284), бараг зөвхөн Британийн хүн амд олддог бөгөөд энэ нь Британийн арлуудад оршин тогтнож байсан урт түүхийг илтгэнэ. Сонирхолтой нь, haplogroup-ийн бага давтамжтай I1Тэгээд I2bЭдгээр нь орчин үеийн Туркийн Битиниа, Галати зэрэг түүхэн бүс нутгуудаас олддог бөгөөд тэдгээрийг Битинийн Никомед I-ийн урилгаар тэнд нүүж ирсэн Кельтүүд авчирсан байж болох юм. Хаплогрупп I2b1Сардины хүн амын ойролцоогоор 1% -д тохиолддог. гэж таамаглаж байна I2b-аас ялгардаг I2Төв Европт, 13 мянган жилийн өмнө аажмаар ухарч буй мөсөн голын ирмэг дээр I2b1-аас I2bбүр хойд зүгт, одоогийн Германы нутаг дэвсгэрт 9 мянга орчим жилийн өмнө. Тодруулбал, Британийн салбар I2b1a-аас ялгарч байв I2b1ойролцоогоор 3 мянган жилийн өмнө. I2b2Хаплогрупп I2b2 нь Германы төв хэсэгт орших хүрэл зэвсгийн үеийн археологийн дурсгал болох Лихтенштейн агуйгаас олдсон араг ясны үлдэгдлээс олдсон бөгөөд тэндээс Урн талбайн соёлын олдворууд олджээ. Агуйд байгаа 19 эрийн үлдэгдлийн 13-аас I2b2 гаплогрупп, нэгээс R1b, хоёроос R1a бүлэг олджээ. Энэ агуй нь I2b хаплогруппын тархалтын голомтод байсан гэж таамаглаж байна. Хаплогрупп I2 нь хуучин Югослав, Румын, Болгар, Сардинид хамгийн түгээмэл эцгийн удам бөгөөд ихэнх славян орнуудад гол удам угсаа юм. Хамгийн их давтамж нь Босни (55%, түүний дотор Боснийн Хорватуудад 71%), Сардини (39.5%), Хорват (38%), Серби (33%), Монтенегро (31%), Румын (28%), Молдав улсад ажиглагдаж байна. (24%), Македон (24%), Словени (22%), Болгар (22%), Беларусь (18.5%), Унгар (18%), Словак (17.5%), Украин (13.5%), Албани ( 13.5%). Энэ нь Германы орнуудад 5-10% давтамжтай байдаг. I2-ийн филогенезХэрэв та генетикийн удмын бичигт шинээр орсон бол филогенетикийн модыг хэрхэн уншихыг ойлгохын тулд манай Филогенетикийн танилцуулгыг уншина уу. I2-ийн филогенетик мод сүүлийн 10 жилийн хугацаанд маш их хувьсан өөрчлөгдөж, дээд түвшний дэд бүлгүүдийн нэрийг R1b-ээс бусад бүх гаплог бүлгийнхээс илүү олон удаа өөрчилсөн. Бүх төөрөгдлөөс зайлсхийхийн тулд хуучин судалгаануудыг унших эсвэл тодорхой дэд бүлэгт ишлэл хийхдээ тодорхойлох мутацуудыг (SNP"s) дахин шалгахыг зөвлөж байна. Хэд хэдэн жижиг дэд бүлгүүдийг орон зайгүйн улмаас оруулаагүй болно.  Томруулахын тулд товшино уу
Гарал үүсэл ба түүхХаплогрупп I бол Европ дахь хамгийн эртний томоохон гаплогрупп бөгөөд тэндээс үүссэн цорын ганц (C1a2 гэх мэт маш бага гаплогруппууд болон бусад гаплогруппуудын гүн дэд бүлгүүдийг эс тооцвол). IJ Haplogroup Ойрхи Дорнодоос Европ руу 35,000 жилийн өмнө ирж, дараа нь IJ haplogroup болон хөгжсөн. Авриньякийн үед (45,000-28,000 жилийн өмнө) Европыг колоничлох анхны хомо сапиенс нь CT, C1a, C1b, F, I гаплогруппуудад харьяалагддаг байсан нь одоо батлагдсан. I2 (M438/P215/S31) нь хожуу палеолитын үед буюу ойролцоогоор 26,500-аас 19,000 жилийн өмнө үргэлжилсэн сүүлчийн мөстлөгийн дээд үе (LGM) үед үүссэн гэж үздэг. Палеолитын үеийн европчууд нүүдэлчин анчин цуглуулагч байсан тул яг гарал үүслийг нь тогтоох боломжгүй ч I2 баруун Европт гарч ирсэн байж магадгүй. Археологийн араг яснаас олдсон хамгийн эртний I2 дээж бол Азилийн соёлтой холбоотой Швейцарийн Гротте ду Бичоноос 13500 настай эрэгтэй хүн юм (Jones et al. (2015)-ыг үзнэ үү). Түүний эхийн удам нь U5b1h байсан. 2016 оны 10-р сарын байдлаар Европын мезолитийн Y-ДНХ-ийн 15 дээжийг туршсан. Тэдний нэг нь C1a2 (Испани), нэг нь F (Германд), хоёр нь I* (Франц), зургаагаас нь I2 (Люксембург, Швед), үүнд I2a1* (P37.2), I2a1a1a багтдаг. (L672), I2a1b (M423) болон I2c2 (PF3827). Тэдний авч явсан эхийн (mtDNA) удам нь U2e, U4, U5a1, U5a2, U5b байв. Оросоос ирсэн дөрвөн дээж нь Y-haplogroup J*, R1a1 (2x) болон R1b1a-д харьяалагддаг. Энэ нь мезолитийн Европын удамшлын дунд тодорхой хэмжээний олон янз байдал аль хэдийн байсаар байгааг харуулж байгаа боловч эдгээр удамшлын ихэнх нь (C1a2, F, I*, J*) одоо маш ховор болжээ. Haplogroup I2a1 нь хараахан тодорхойгүй байгаа шалтгааны улмаас неолитын үеэс том ялагч болж гарч ирсэн бололтой. Матиесон нар Анатолиас ирсэн неолитын эхэн үеийн геномын шинжилгээ. (2015) I2c (L596) нь 8500 жилийн өмнө Туркийн баруун хойд хэсэгт аль хэдийн байсан бөгөөд үржил шимт хавирган сараас Европ руу явах замдаа неолитын үеийн фермерүүд түүнийг уусгаж байсныг харуулж байна. 2014-2016 оны хооронд явуулсан янз бүрийн судалгаагаар мезолитийн үеийн Европын бүх удам шинэ чулуун зэвсгийн үеийн тариачдын давалгаанд шингэсэн болохыг тогтоожээ. Гэсэн хэдий ч I2a нь тэдний дунд хамгийн түгээмэл нь байсан бөгөөд туршилтанд хамрагдсан неолитын Y-ДНХ-ийн 69 дээжийн 16% -ийг эзэлдэг. Энэ нь түүнийг Анатолийн тариачдын анхны удам угсаа болох G2a-ын дараа хамгийн түгээмэл хоёр дахь эцгийн удам юм. I2a1 нь ялангуяа Зүүн өмнөд Европ дахь Старчево-Корс-Кришийн соёлд (МЭӨ 6000-4500) цэцэглэн хөгжсөн бололтой, энэ нь өнөөг хүртэл гол эрэгтэй удмын нэг хэвээр байна. I2a1 нь Газар дундын тэнгисийн баруун хэсэгт хэвлэмэл кардиум ваарны соёлд (МЭӨ 5000-1500) мөн гарч ирсэн. Орчин үеийн Сардинчууд болон Баскууд өөрсдийн геномын хамгийн их хувийг Неолитын үеийн фермерүүдээс хэвлэмэл-кардиум ваарны соёлоос өвлөн авсан. Хэдийгээр неолитын эхэн болон дунд үеийн дээжээс өнөөг хүртэл ганцхан I2a2 (Испаниас авсан I2a2a-M223) олдсон боловч тэдгээрийн ихэнх нь зэс, хүрэл зэвсгийн үед илэрчээ. Зэс ба хүрэл зэвсгийн үеийн дээж нь Испанид нэг I2a2, хоёр I2a2a, нэг I2a2a1 (CTS616), Оросын өмнөд хэсэгт I2a2a1b1b2 (S12195), нэг I2a2a1 (CTS9183), нэг I2a2a1b1b2 (S12195) (CTS9183). ), Германд зургаан I2a2b (L38) (Unetice болон Urnfield соёл). Зэсийн үеийн I2a1 дээжүүдэд хойд Италийн I2a1a1 (Ремеделло соёл), Унгараас гаралтай I2a1 (Ватя соёл) багтдаг. I2c2 нь Герман дахь Юнетикийн соёлд мөн олдсон. Haplogroup I2a1 (P37.2)Haplogroup I2a1 нь I2 бүлгийн хамгийн том салбар бөгөөд зүүн өмнөд, баруун өмнөд болон баруун хойд Европ дахь неолитын соёлтой хамгийн хүчтэй холбоотой нэг юм.  Haplogroup I2a1a1 (M26)I2a1a (М26, L158, L159.1/S169.1) нь 2005 он хүртэл I1b2, 2006-7 онд I1b1b, 2008-2010 онд I2a1 гэж нэрлэгддэг байсан. Энэ нь Баруун Европ даяар тархсан бөгөөд Сардинчуудын дунд хамгийн их давтамжтай (37.5%) хүрдэг. ба Баскууд (5%), хоёр хүн амыг тусгаарладаг. M26 нь газарзүйн хувьд Британийн арлууд, нам дор улсууд, Франц, баруун Герман, Швейцарь, Сардини, Сицили, Италийн баруун эрэг, Ибериа, Магребын Газар дундын тэнгисийн эрэгт хязгаарлагддаг. L160 мутацийн цорын ганц M26 сөрөг нь Ирландад л байдаг. I2a1a-M26 нь неолит ба халколитийн үеийн баруун Европын мегалитийн соёлын үндсэн эцэг эхийн нэг байсан байх. I2a1a1a (L672) нь мезолитийн үеийн Шведэд аль хэдийн олдсон бөгөөд энэ нь мезолитийн үед I2a1a нь Ибериас Скандинав хүртэл маш өргөн тархсан байсан гэсэн үг юм. Хожим нь тэд Ойрхи Дорнодын шинэ хүмүүстэй холилдон газар тариалан эрхлэх байсан. Haplogroup I2a1b (M423)I2a1b (M423, L178) нь 2007 он хүртэл I1b, 2008-2010 он хүртэл I2a2 гэж нэрлэгддэг байсан. Бүх M423 удмын 90 гаруй хувийг төлөөлдөг үндсэн дэд бүлэг нь L621ба түүний дэд бүлэг L147.2. Бусад дэд ангиуд нь L41.2(маш ховор) ба L161.1(Герман болон Британийн арлуудад ихэвчлэн олддог). Haplogroup I2a1b-L621Энэ салбар нь ихэвчлэн Славян орнуудад байдаг. Түүний хамгийн их давтамж нь Динар Славууд (Словен, Хорват, Босниак, Серб, Монтенегро, Македон) болон Болгар, Румын, Молдав, баруун Украин, Беларусь зэрэгт ажиглагддаг. Мөн Албани, Грек, Унгар, Словак, Польш, Оросын баруун өмнөд хэсэгт бага хэмжээгээр түгээмэл байдаг. I2-L621 (L147.2+) гэж бас нэрлэдэг I2a-Din(Динарикийн хувьд). Зүүн хойд Румын, Молдав, Украины төв хэсэгт I2a1b-L621-ийн өндөр концентраци нь Кукутени-Трипиллийн соёл (МЭӨ 4800-3000) хамгийн их тархсаныг сануулж байна. Энэ соёлын Y-ДНХ-ийн дээжийг өнөөдрийг хүртэл туршиж үзээгүй боловч Старчево-Корс-Крисийн соёлын нэг салбар болж хөгжсөн тул I2a нь түүний эцгийн удмын нэг байсан байх магадлалтай бөгөөд үүсгэн байгуулагчийн нөлөө нэмэгдэж магадгүй юм. түүний давтамж Кукутени-Трипиллийн соёл нь хүрэл зэвсгийн үеийн Энэтхэг-Европын довтолгооноос өмнө Европ дахь шинэ чулуун зэвсгийн үеийн хамгийн дэвшилтэт соёл байсан бөгөөд Ямна хотыг Балкан болон Төв Европ руу тэлэхээс өмнө тал нутгийн соёлтой эрчимтэй холбоотой байсан бололтой (R1a-ийн түүхийг үзнэ үү). ба R1b). МЭӨ 3500 оноос хойш Понти-Каспийн тал нутагт Ямнагийн үе эхлэхэд Кукутени-Трипиллианчууд зүүн тийш тэлсээр одоогийн баруун Украины тал руу орж, хотуудаа (тухайн үеийн дэлхийн хамгийн том) орхин гарчээ. мөн Ямнагийн хөршүүд шиг улам бүр нүүдэлчин амьдралын хэв маягийг баримталж байна. Кукутени-Трипиллианчууд Ямнагийн хөршүүдэд уусаж, утаснуудын тэлэлтээр Балтийн эрэг рүү урагшлах явцдаа R1a, R1b гаплогруппуудтай зэрэгцэн цөөнхийн удам болон тархсан гэж амархан төсөөлж болно. Өөрөөр хэлбэл, I2-L621 удам угсаа нь Украйн, Польш эсвэл Беларусийн хаа нэгтээ орших Прото-Индо-Европын нийгмээс харьцангуй тусгаарлагдмал амьдарч болох байсан бөгөөд дараа нь олон зуун, мянган жил өнгөрөх тусам тэдний эргэн тойрон дахь голчлон R1a популяцитай холилдох байсан. Үүссэн амальгам нь Прото-Славуудын өвөг дээдэс болох байсан. Өнөө үед I2a1 нь Зүүн өмнөд Европт G2a-аас 5-10 дахин их байдаг бол неолитын үед G2a ойролцоогоор 4 дахин их тархсан байна. Энэ бүрэн урвуу байдлыг юу тайлбарлаж болох вэ? Түүхийн нэгэн цагт I2a1 удмынхан ялалт байгуулсан талдаа ашигтай байсан юм шиг санагддаг. Ямна баруун тийш Европ руу тэлэхэд бага зэрэг түлхэц үзүүлснээс гадна I2a1b-L621-ийг Зүүн Европын гол удам болох боломжийг олгосон гол шийдвэрлэх үйл явдал нь МЭ 6-9-р зууны хооронд славянчуудын нүүдэл байсан байх магадлалтай. Орчин үеийн Зүүн Европын ихэнх хүмүүс I2a1b-д хамаарах L147.2 (CTS10228, CTS2180 эсвэл Y3111) дэд бүлэгт багтах бөгөөд энэ нь 5600 жилийн өмнө үүссэн гэж үздэг (Ямнагийн үе ба Трипиллийн хээр тал руу тэлэхийн өмнөхөн) боловч TMRCA нь ердөө 230. Yfull-ийн мэдээлснээр L147.2-д сөрөг I2a1b-L621 бодгальуудын цөөнх нь Польшийн зүүн хэсэг, Беларусь, баруун Украины орчимд байдаг бөгөөд энэ нь хальколитын үеэс хойш энэ удам оршин тогтнож ирсэн болохыг харуулж байна. I2a1b-L147.2 дэд бүлэг 1900 жилийн өмнөхөөс маш хурдацтай өргөжсөн нь славян угсаатны нийлэгжилтийн цаг үетэй нийцэж байгаа бөгөөд нэг хүн хүн амын хэмжээнд хангалттай хэмжээний эрэгтэй үр удамтай болох хүртэл хэдэн зуун жил шаардлагатай байдаг. Энэхүү I2-L147.2 өвөг дээдэс нь 2300 жилийн өмнө бага байсан ч эрчимтэй хөгжиж буй Эрт Славян хүн амд ийм нөлөө үзүүлэх болно. Гот, Вандал, Бургунд зэрэг зүүн Герман, Польшид амьдардаг герман овог аймгууд Ромын эзэнт гүрнийг довтолсны дараа зүүн талд амьдардаг Славууд орон зайг дүүргэсэн. 476 онд Баруун Ромын эзэнт гүрэн задран унасны дараа славянчууд Динарийн Альп болон Балканы хойг руу нүүжээ. 9-р зуун гэхэд Славууд түрэг хэлээр ярьдаг Булгаруудын мэдэлд байсан зүүн Балканы нутгаас бусад орчин үеийн славян хэлээр ярьдаг бүх нутаг дэвсгэрийг эзэлжээ. Одоогийн байдлаар хойд славян орнуудад I2a-L621-ийн 9% (Польш, Чех) - 21% (Украйн) байдаг бол өмнөд Славууд 20% (Болгар) -аас 50% (Босни) хооронд байна. Өмнө зүгт I2a-Din-ийн өндөр хувь нь өмнөд славянчууд баруун Украинд гаралтай бөгөөд I2a-ийн R1a-ийн харьцаа өндөр байсантай холбоотой өөр нэг үүсгэн байгуулагч нөлөөлөлтэй холбоотой байж магадгүй юм. Бараг бүх Dinaric I2a нь L147.2 салбар, дийлэнх нь ердөө 1800 жилийн өмнө амьдарч байсан нийтлэг өвөг дээдсээс гаралтай S17250 салаа бүлэгт багтдаг. Хаплогрупп I2a1b-L161.1Генетикийн удамшлын хүрээлэлд ихэвчлэн мэддэг I2-M423-Арлууд, L161.1 нь баруун Ирланд (5-10%) болон Шотландын уулархаг нутагт (1-5%) хамгийн өндөр давтамжтай байдаг боловч Төв болон Баруун Европ даяар бага давтамжтай (> 1%), Латви, Литва, Беларусь нь Британийн арлууд хүртэл, Скандинаваас баруун хойд Испани хүртэл. Мөн Албани, Грекийн хойд хэсэг, Болгар, Румынд ч олджээ. Мэдэгдэж байгаа хамгийн эртний I2a1b-L161.1 хүмүүс бол Мезолитын Люксембургийн 8000 настай Лосчбур, Шведийн өмнөд хэсэгт орших Мотала хотын 7800 настай эрэгтэй хүмүүс юм. I2a1b-L161.1 нь мөстлөгийн төгсгөл ба мөстлөгийн дараах үеийн үед Европын ихэнх хэсэгт тархсан байж магадгүй бөгөөд бусад мезолитын үеийн нэгэн адил неолитын нийгмийг нэгтгэсэн байх магадлалтай. I2a1b-L161.1 нь Мегалитийн үед Британийн арлууд дахь неолитын гол удмын нэг байсан тул Стоунхенж, Ноут, Ньюгранж болон бусад агуу Мегалитийн дурсгалт газруудыг барьсан хүмүүсийн дунд байх байсан байх. Өнөөдөр түүний давтамж бага, Европын баруун хойд болон зүүн өмнөд захад хязгаарлагдаж байгаа нь сүүлийн 5000 жилийн хугацаанд Индо-Европын олон тооны нүүдлийн давалгааны үр дүн юм. Proto-Celtic гаплогрупп R1b-L21 одоо МЭӨ 2000 онд Ирландад ирсэн нь мэдэгдэж байна (Cassidy et al. 2015-ыг үзнэ үү), R1b Төв Европт анх ирснээс хойш хэдхэн зууны дараа. Шилжилт хурдацтай явагдаж, R1b овог аймгууд Британийн арлуудын неолитийн үеийн хүн амыг хэдхэн зууны дотор дарж, тэднийг баруун болон хойд зүг рүү түлхэж, одоо L161.1 оршин тогтнож байна. Haplogroups I2-L880, I2-L1286 болон I2-L1294Эдгээр жижиг дэд бүлгүүд нь ихэвчлэн баруун хойд Европт бага давтамжтай байдаг. I2a2 шиг (доороос харна уу) эдгээр удам угсаа нь төв болон баруун хойд Европын анчин цуглуулагчдын үр удам байж магадгүй юм. L880Энэ нь Герман, Люксембург, Францад танигдсан маш ховор дэд бүлэг юм. L1286хожуу палеолитын үед үүссэн. Гэсэн хэдий ч орчин үеийн тээвэрлэгчдийн 99% нь L233 дэд бүлэгт харьяалагддаг бөгөөд 2000-аас бага жилийн өмнө нийтлэг өвөг дээдэстэй байжээ. L233 нь Британийн арлуудад хамгийн түгээмэл боловч Нидерланд, Скандинав, Герман, Франц, Польшид хааяа тааралддаг. L1294Мөн маш ховор бөгөөд зөвхөн Герман, Франц, Британид л олдсон. Haplogroup I2a2 (P214)I2a2 (S33/M436/P214, P216/S30, P217/S23, P218/S32, L35/S150, L37/S153, L181) 2005 он хүртэл I1c, 2010 он хүртэл I2b гэж нэрлэгддэг байсан. өмнөх Кельт-Германзэрэг баруун хойд Европын хүмүүс мегалит барилгачид(МЭӨ 5000-1200). Түүний насыг 21,000-аас 13,000 жилийн хооронд тооцоолсон бөгөөд энэ нь эпипалеолитын үетэй тохирч байна. I2a2 нь Европын ихэнх хэсэгт байдаг бөгөөд неолитын үеийн фермерүүд ирэхээс өмнө тив даяар тархсан бололтой. Өнөөдрийг хүртэл хэд хэдэн мезолитийн I2a2 дээжийг тодорхойлсон бөгөөд ихэнхдээ Матиесон нар. (2017) Үүнд өмнөд Герман (МЭӨ 7200 оны M223), Серби, Румыны хоорондох Төмөр хаалга (МЭӨ 6200 оны үеэс Z161), Латви (МЭӨ 5500 оны үеэс CTS10057) болон зүүн өмнөд Украин (МЭӨ 5400 оны үеэс L699) зэрэг хүмүүс багтана. болон МЭӨ 5200 он хүртэл L701). Нэмж дурдахад дэд бүлэг I2c1 (L1251)саяхан тогтоогдсон. Энэ нь ихэвчлэн Герман, Англид болон тэдгээрийн захад (Ирланд, Норвеги, Франц, Итали, Польш) байдаг. Энэ дэд бүлэг нь ойролцоогоор 6000 жилийн настай гэж үздэг бөгөөд энэ нь түүнийг сүүлийн неолитын үед байрлуулдаг. Энэ нь Шугаман вааран эдлэлийн соёлоос гаралтай Ойрхи Дорнодын тариачидтай холбоо тогтоосны дараа газар тариалан эрхэлсэн Германаас ирсэн анчин цуглуулагчидтай холбоотой байж магадгүй юм. Алдартай хүмүүсI2a1b-L621 салбар (Славян)I2a2a-Z161 салбар (Герман)Эндрю Жонсон(1808-1875), АНУ-ын 17 дахь Ерөнхийлөгч, 16 дахь дэд ерөнхийлөгч нь I2a2a (хуучин I2b1) хаплогруппын гишүүн болохыг судалгааны үр дүнд үндэслэн тогтоожээ. Хэвлэлд Эхлээд I2a1 гаплогруппийг авч үзье (мутаци P37.2). Славууд (мөн бараг зөвхөн Славууд!) нь түүний салбар I2a1b (мутаци L178/S32 ба M423), эсвэл илүү нарийн, дэд салбар I2a1b3 (мутаци L621/S392) болон түүний охин I2a1b3a (L147.2) -аар тодорхойлогддог. L621/S392 болон L147.2 мутаци байгаа эсэхийг маш цөөн тооны хүмүүс туршиж үзсэн тул илүү нарийн хэлэхэд хэцүү хэвээр байна. Славуудаас гадна тэдний дунд Герман, Итали, Их Британийн тусгаарлагдсан төлөөлөгчид байдаг. Y хромосом дахь 58 сая нуклеотидын дунд славян дэд салбараас бүр ч хожуу мутаци илрэх магадлалтай. I2a1 хаплогруппын славян дэд салбарын тархалтыг хоёртын тэмдэглэгээгээр бус харин түүний онцлог гаплотипээр илүү сайн ажигладаг. I2a1b дэд салбарт тэдгээр нь маш өвөрмөц бөгөөд найдвартай төлөөлөгчдөө тодорхойлох боломжийг олгодог. ДНХ-ийн удам судрыг сонирхож буй хүмүүст зориулсан арилжааны шинжилгээ хийдэг FamilyTreeDNA (FTDNA) мэдээллийн сан нь ийм туршилтын явцад тодорхойлсон бүх гаплотипуудыг агуулдаг. Бидний сонирхож буй дэд салбарыг тэнд "Динарик кластер" гэж тодорхойлсон. Энэ нь бүх славян популяцид төлөөлдөг өмнөд, үнэндээ динар, хойд гэсэн хоёр салбар руу хуваагддаг (үүнийг "Карпатын" гэж нэрлэх нь илүү зөв байх болно). Энэхүү мэдээллийн санг ашиглан янз бүрийн популяци дахь салбаруудын давтамжийг тодорхойлох боломжгүй, учир нь энд үзүүлсэн дээжүүд нь санамсаргүй байдлаар үүсдэг (ДНХ-ийн шинжилгээ хийлгэх хүсэл, боломжийн давхцал), хэмжээ нь маш өөр бөгөөд төлөөлдөггүй. нийт хүн амын тоо. Арга зүйн хувьд зөв дээжийг шүүх эмнэлгийн зорилгоор бий болгож, хадгалсан Y-гаплотипуудын (YHRD) олон улсын лавлагааны мэдээллийн сангаас, мөн Беларусийн Бүгд Найрамдах Улсын хүн амын ДНХ маркеруудын ижил төстэй мэдээллийн сангаас олж болно. Бүгд Найрамдах Беларусь улсын Хууль зүйн яамны Шүүхийн шинжээч, криминалистикийн төвөөс бүтээсэн [Беларусь дахь Y-STR локалуудын гаплотипуудын давтамжийн лавлагаа мэдээллийн сан: http://dnkexpertiza.org/index1.html ]. YHRD-д ороогүй зарим дээжийн хаплотипийг шинжлэх ухааны хэвлэлээс олж болно. Эдгээр бүх дээжийг FTDNA-тай харьцуулахад харьцангуй цөөн тооны локусуудаар төлөөлдөг: арилжааны туршилтууд нь 37, 67, бүр 111 локусын багцыг ашигладаг бөгөөд шүүх эмнэлгийн шинжилгээнд Y-Filer гэж нэрлэгддэг 17 цэгийн багцыг ашигладаг. Стандарт. Гэхдээ энэ багц нь ч гэсэн Динарик кластерт хамаарах гаплотипуудыг итгэлтэйгээр таних боломжийг олгодог. Эдгээр нь энэ кластерын модаль гаплотипээс харьцангуй бага генетикийн зайгаар тодорхойлогддог бөгөөд үүнийг 1-р хүснэгтэд Y-Filer форматаар харуулав. Хүснэгт 1. I2a1 гаплогруппын Динар кластерын модаль гаплотип
Энэ профайлаас нийтдээ 9-10 алхамаар (давталтын тооны мутаци) ялгаатай хаплотипууд нь ихэнх тохиолдолд бидний сонирхдог I2a1 салбарт хамаардаг. Тэдгээрийг тодорхойлоход гарч болзошгүй алдаа нь 1-2% -иас хэтрэхгүй бөгөөд энэ нь дээжийг хоёртын тэмдэглэгээг ашиглан нэгэн зэрэг туршсан тохиолдолд (харамсалтай нь цөөхөн) нотлогддог. Динарик кластерын өмнөд салбар нь 67 ба түүнээс дээш 111 локустай нэлээд сайн ялгардаг боловч 17-локустай Y-Filer багцад цорын ганц ялгагдах онцлог нь DYS-448 локусын давталтын тоог багасгасан явдал юм: 19 ( Заримдаа бүр 18) 20-ийн оронд. Мэдээжийн хэрэг, хоёр дэд кластерийг зөвхөн нэг локусыг ялгах нь зөвхөн эхний ойролцоолсон байдлаар боломжтой боловч ерөнхий дүр зураг ийм бүдүүлэг тооцоололд ч тодорхой байна. Y-Filer форматын нийтлэгдсэн бүх дээжийн хувьд энэ кластерт хамаарах гаплотипуудын давтамжийг 2-р хүснэгтэд үзүүлэв. Хүснэгт 2. Динар кластерын гаплотипуудын давтамж
Хүснэгтээс харахад I2a1 гаплогруппын динар кластер нь Балкан-Карпатын бүс нутагт, Баруун болон Зүүн Славуудын дунд Лука-Райковецын соёлын бүсэд хамгийн их байдаг. Польш, Орос хоёрын аль алинд нь түүний давтамж буурч, Литва, Германд энэ нь славян хольцтой сайн тайлбарлагдахуйц түвшинд байдаг. Энэ нь Лусат-Помераны нутгаас прото-славян хэлийг авчирсан Бастарни, Венетийн аль алинд нь энэ бөөгнөрөл, түүнчлэн славянуудтай эрчимтэй холбоо тогтоож эхлэхээс өмнө эртний Балтчууд байхгүй байсан гэсэн үг юм. Автохтон популяцийн нэлээд хэсэг нь ууссаны үр дүнд орчин үеийн славянчуудын удмын санд оржээ. Славян бус бүх ард түмнээс I2a1 хаплогрупп нь зөвхөн Румынчуудын дунд өндөр давтамжтай байдаг бөгөөд энэ нь эртний славянууд (Прага ба Ипотести-Чурлийн соёлыг тээгчид) Фракийн үр удам болох Влахчуудтай холилдсоны үр дүнд бий болсон. хүн ам Ромын эзэнт гүрний үед романчлагдсан. Түүгээр ч зогсохгүй румынчуудын дунд болон славян угсаатны бүлэг (Львов муж, Беларусийн өмнөд хэсэг) үүсэх давтамж ойролцоогоор ижил байна. Тиймээс Влахчуудын дунд энэ нь эхэн үеийн Славуудынхаас доогуур байсангүй. Өмнөд дэд кластерын тархалт нь шинж тэмдэг юм. Энэ нь ерөнхийдөө славян популяцийн онцлог шинжгүй юм. Зүүн Славян ба Польшийн дээжид байдаг DYS448=19-тэй цөөн хэдэн гаплотипууд нь үлдсэн локусуудын модаль утгуудаас нэлээд хол бөгөөд бие даасан мутацийг тусгасан байж магадгүй юм. Үүний зэрэгцээ, Өмнөд Славян гаплотипууд нь ихэвчлэн модальтай маш ойрхон байдаг. Тэдний ихэнх тээгч нь DYS448 байрлалд 19 удаа давтагдсан ганц хүний үр удам болох нь тодорхой. Энэ хувь хүн бүх Динар кластерын өвөг дээдсээс хожуу амьдарч байсан боловч тийм ч их биш - ерөнхийдөө өмнөд дэд кластерт гаплотипуудын тархалт нэлээд том, өвөг дээдсийн гаплотипээс 6-7 алхмын зайтай генетикийн зай нь тийм ч ховор биш юм. Хүрэл зэвсгийн төгсгөл - Төмөр зэвсгийн эхэн үед 20 давталттай өвөг дээдсийн бөөгнөрөл, 19 давталттай охин дэд кластер хоёулаа Карпатын өмнөд хэсэгт тархсан бололтой. нутаг дэвсгэрийн нэг хэсэг байсан бөгөөд славянууд гарч ирэхээс өмнө Карпатын сав газарт бүхэлд нь тархаж амжаагүй байв. МЭ 271 онд Дачиаг нүүлгэн шилжүүлсний дараа. д. Энэ нь зөвхөн Дунай мөрнийг бараг гаталж үзээгүй Византийн харьяат хүмүүсийн онцлог шинж байсан бололтой. Үүний зэрэгцээ, DYS448=20 бүхий хуучин бөгөөд илүү өргөн хүрээтэй дэд бүлэг нь Карпатын хойд хэсэгт нэвтэрч, шинээр гарч ирж буй Славуудын нэг хэсэг болжээ. Славуудын өвөг дээдэс "Динарик" гаплотипийн өндөр хувьтай бүлгийг хэзээ, хаана оруулсан бэ? Энэ нь 2-р зуунаас өмнө тохиолдож болохгүй. МЭӨ. - Зарубинцы соёл үүссэн үе. Гэхдээ энэ нь хойд зүг рүү буцах нүүдэллэх явцад Лука-Райковецын соёл бий болсон 7-8-р зууны эхэн үеэс хойш тохиолдож болохгүй. Энэ гаплогрупп байхгүй байсан нутаг дэвсгэрт үүссэн Орос, Польшчуудын дунд орчин үеийн I2a1 давтамжийг тайлбарлах өөр арга байхгүй. Энд хоёр боломжит хувилбар байна. I2a1-ийн тээвэрлэгчид нь Зарубинцын соёлыг бий болгоход оролцсон нутгийн соёлын аль нэг эсвэл хожуу Зарубинцын бүлгүүдийн аль нэгний төлөөлөгчид байж болно. Бид Милоград-Нейрүүд, Борисфенитүүд-скифүүд эсвэл Юхновитууд-Меланхленчуудын тухай ярьж болно. Хоёрдахь хувилбарт удамшлын сангийн энэ бүрэлдэхүүн хэсэг нь шинээр гарч ирж буй славянчуудын суурьшлын явцад, Украин дахь Черняховын соёлын үлдэгдлийг уусгах явцад эсвэл Карпатын сав газар, Дундад Дунай дахь Романжуулсан Фракийн үндэстнийг шингээх явцад олж авсан. Зарубинцы соёлд (ялангуяа Дээд Днеприйн хувилбарт) Милоградын субстрат байгаа нь нэлээд магадлалтай боловч янз бүрийн судлаачдын түүний үүргийн талаарх үнэлгээ өөр өөр байдаг. Хожуу Скифийн субстрат нь Дундад Днепр хувилбарт тодорхой харагдаж байна. Хожуу Зарубинцы бүлгүүд болох Почеп, Картамышев, Грини-Абидня зэрэг бүлгүүдийг байгуулахад Юхновскийн чухал бүрэлдэхүүн хэсэг оролцсон боловч сүүлийнх нь Киевийн соёлыг бий болгоход шийдвэрлэх үүрэг гүйцэтгэсэн. Гэхдээ энэ хувилбар нь хойд болон өмнөд дэд кластеруудын гаплотипуудын ижил төстэй байдлыг муу тайлбарлаж байна. Юхновын соёлын гарал үүслийн талаар мэддэг зүйл нь Балкан-Карпатын бүс нутгийн соёлд нийтлэг байдаг бүрэлдэхүүн хэсэг байгаа талаар ярих боломжийг бидэнд олгодоггүй. Милоград ба ойт хээрийн скифийн соёлд ийм бүрэлдэхүүн хэсэг нь МЭӨ 1-р мянганы эхэн үед Чернолесийн соёл үүсэх үед тэмдэглэгдсэн баруун өмнөд зүгийн тодорхой түлхэц байж болно. д. Прото-Фракийн тамгатай ба лимбэтэй керамик бүтээгчидтэй холбоотой популяци нь "Динарик" гаплотипуудыг Украйны ойт хээрт авчирч, дараа нь скиф, Милоградианчуудад өв залгамжлалаар дамжуулж болох байсан. Гэхдээ эдгээр гаплотипуудын тээгч нь олон тооны тодорхой мутаци хуримтлуулж, Фракийн үндсэн ертөнцөөс тусгаарлагдсан хэдэн зууны турш оршин тогтнож байсан байх ёстой. Үүний зэрэгцээ Беларусь, ОХУ-ын нутаг дэвсгэр дээр I2a1 гаплотипуудад онцгой өвөрмөц байдал байдаггүй. Хоёрдахь хувилбарт Хойд Динарийн гаплотипуудын донорууд нь тэлэлтийн явцад Прагийн соёлд шингэсэн Карпатын довны соёл (энэ нь Карп овогтой тодорхойлогддог) ба Дако-Ромын үлдэгдэл бүлгүүд юм. Карпатын сав газрын хүн ам. Славууд Авар хаант улсын дотор тэдэнтэй холилдох үед хойд хэсгийн довтолгооны давтамж 20 орчим хувьтай байв. Дараа нь тэд гаплотипуудаа ойн бүсэд хэсэгчлэн зөөвөрлөж, нутгийн хүн амтай холилдох тусам давтамж буурчээ. Цаашид өмнө зүгт Византийн эзэнт гүрэн рүү явсан хүмүүс тэнд Өмнөд Динарийн гаплотип ихтэй автохтонуудтай тааралдав. Хорватууд, сербүүд болон хэсэгчлэн Македончууд ерөнхийдөө I2a1-ийн өндөр давтамжийг өртэй бөгөөд энэ нь ерөнхий славян түвшнээс хамаагүй давсан байдаг. Дунайгаас өмнө зүгт суурьшсан славянчууд буцах нүүдэлд бараг оролцдоггүй байсан тул ойн бүсэд Өмнөд Динарийн гаплотип цөөхөн байсан. Эдгээр гаплотипуудыг Дунай мөрний хойд хэсэгт бөөнөөр нь авчирсан цорын ганц хүмүүс бол Австричуудын славян өвөг дээдэс, түүнчлэн Румынчуудын өвөг дээдэс болох Влахчууд байв. Y-хромосомын гаплогрупп I2a-ийн гарал үүсэл, түүний хувьсал, энэ гаплогруппын тээвэрлэгчдийн популяцийн шилжин суурьших зам нь хүний популяцийг идэвхтэй судлах сонирхолтой болсноос хойш 5 жилийн турш миний тусгай судалгааны сэдэв байсаар ирсэн. генетик. Миний энэ гаплогруппыг сонирхож байгаа шалтгаан нь маш энгийн - миний Y-хромосомын генетикийн шинжилгээ (Y-STR + Y-snips) нь филогенетикийн ангиллын үүднээс энэ нь энэ төрлийн нэг салбартай болохыг харуулсан. хаплогрупп, мөн энэхүү улиг болсон биологийн баримтын улмаас би энэ гаплогруппын эртний төлөөлөгчдөөс шууд эрэгтэй шугамаас гаралтай. Энэ гаплогруппын судалгаанд миний оролцоо багатай чухал шалтгаан бол I2 болон I-ийн бүх хаплогруппын талаар харамсалтай мэдээлэл дутмаг байгаа явдал юм. Энэ гаплогруппын талаарх хамгийн сүүлийн үеийн тусгай судалгаанууд 2004, 2006, 2007 онд хэвлэгдсэн гэдгийг хэлэхэд хангалттай. - Rootsi , Сиири гэх мэт.(2004), Rootsi, S. (2006), Peter Underhill нар (2007). Үлдсэн бүтээлүүд, ялангуяа Балкан болон хуучин Югослав дахь популяцийн Y-хромосомын олон янз байдлын талаархи бүтээлүүд нь дээрх гурван бүтээлд дурдсан дүгнэлтийг давтаж, өөр өөр зүйл нэмэлгүй (Маржанович, Дамир; et al. 2005, Peričić) , Марижана гэх мэт. 2005; Ребала К., гэх мэт.(2007). Тиймээс Эстонийн попгенетик судлаач Роотсигийн бүтээл I гаплогруппын гарал үүслийн талаархи хэд хэдэн үндсэн таамаглалыг тодорхойлсон бөгөөд хожим нь шинжлэх ухаан, алдартай шинжлэх ухааны (Википедиа) эх сурвалжуудад нэгтгэгджээ. Харамсалтай нь эдгээр таамаглалуудын ихэнх нь худал болж хувирсан. Доор бид Википедиагийн I2-ийн тухай нийтлэлд агуулагдах зарим заалтыг албан ёсны консерватив популяцийн генетикт хүлээн зөвшөөрдөг бөгөөд нийтлэлээс харахад дээрх заалтуудын заримыг хожим нь сонирхогчдын генетикийн ойрын хүрээлэлд дахин бодож, боловсруулсан болно.
Гаплогрупп ба хэл, гаплогрупп ба археологийн соёлын хоорондын уялдаа холбоо (эдгээр холболтыг тусад нь судлах нь зүйтэй) гэсэн асуултыг орхиж, бид Балканы өвөг дээдсийн төрөлх нутаг I-ийн талд Роотсигийн гол аргумент нь Динар хэлэнд энэ гаплогруппын чухал ач холбогдолтой байдгийг олж харах болно. Альп/Балканы бүс нутаг. Ийм тоонуудын хамгийн энгийн бөгөөд хямдхан тайлбар нь Европт (20 мянган жилийн өмнө) хамгийн сүүлчийн мөстлөгийн үеэс хойш Балканы хүн амын удмын сангийн нэг хэсэг нь тасралтгүй үргэлжлэх болно гэж үзэх явдал юм. Балканы мөстлөгийн хоргодох үеэс хойш. Мэдэгдэж байгаагаар Y-хромосомын гаплогруппуудын газарзүйн гарал үүслийг дараахь эмпирик шалгуураар тодорхойлж болно: тухайн гаплогрупп үүссэн газар эсвэл популяцид, түүний давтамж ба STR тархалт (эсвэл STR хувьсах нас)бусад популяцитай харьцуулахад хамгийн их байна (Sengupta et al., 2006). Энэхүү энгийн бөгөөд логик мэт санагдах схемийг I haplogroup-ийн жишээн дээр илүү нарийвчлан авч үзэхэд шинэ өгөгдлийн үүднээс шүүмжлэлийг тэсвэрлэдэггүй. Гэсэн хэдий ч, Роотсигийн нийтлэлүүдийн нөхцөл байдал нь генетикийн эрдэмтэд гаплогрупп I-ийн бүтцийн талаар ерөнхий ойлголттой байсан тэр үед хийгдсэн гэдгийг санах нь зүйтэй. Популяцийн төрөлжсөн генетикч нарын дараагийн судалгаа, генези дэх гаплогрупп I-ийн тусгай сонирхогчдын судалгаатай хослуулсан. арилжааны төслүүд FTDNA (Family Tree DNA - Haplogroup I Y-DNA Project, FTDNA I1 yDNA Haplogroup, FTDNA I2*New ISOGG I2b, FTDNA I2a Y-Haplogroup, FTDNA I-M223 Y-Haplogroup, FTDNAL8, ялангуяа ихээхэн хүчин чармайлт) ДНХ-ийн угийн бичиг гэж нэрлэгддэг сонирхогч судлаачдын (Кеннет Нордтведт, Берни Каллен, Стив Трангсруд, Уильям Хартли, Име Дрост, Аарон Ж. Хилл, Питер Хречдакян, Хованн Симониан, Боб Мэй, Вадим Веренич, Билл Морроу, Лоуренс Майка, Владимир Семарг) гэх мэт) нь хаплогруппын салбаруудын филогенетик харилцаа, бүтцийг илүү сайн, илүү сайн ойлгох боломжийг олгож, хаплогруппын салангид салбар-дэд бүлэгт зориулсан он дараалал, газарзүйн лавлагаа өгөх боломжийг олгосон. Энэхүү судалгаа нь I2a хаплогруппын гарал үүсэл, "хувьсал"-ын талаарх бидний мэдлэгийн өнөөгийн байдлын талаар товч мэдээлэл өгөхөөр хязгаарлагдах болно. I2a хаплогруппын гарал үүсэл, шилжих замууд. Шүүмжийн хэсэгчилсэн шинж чанараас шалтгаалан би хаплогрупп Европт хэрхэн, хэзээ үүссэн тухай ижил чухал асуултууд, мөн I-ийн суурь салбарыг I1 ба I2 гаплогруппуудад хуваах цаг хугацаа, газартай холбоотой асуултуудыг хөндөхгүй. Одоогийн байдлаар бидэнд хувилбарын олон тооны, адил магадлалтай хувилбаруудын аль нэгийг батлах хангалттай нотлох баримт байхгүй тул бид Дундад палеолитын Европын оршин суугчдын ДНХ-ийн судалгааг удахгүй хийх найдвараар эдгээр асуултыг зүгээр л орхих болно. Үүний оронд бид I2a гаплогруппын филогенетик ('гэр бүлийн мод') модыг танилцуулж, энэ бүлгийн бусад салбаруудыг орхиж тайлбарыг эхлүүлнэ. Хаплогрупп I-ийн стандарт энгийн диаграммуудын оронд бид Нордтведтийн боловсруулалт дахь модны каноник хувилбарыг санал болгож байна (2013 оны хувилбар). Үүний гол давуу тал нь дэд бүлгийн салбарууд ба гаплотипуудын интраклад кластеруудын харьцангуй топологийн байршлын дүрслэл юм. Нэмж дурдахад, модны доорх масштабыг Кен Нордтведтийн хөндлөн тархалтын аргаар тооцоолсон мөчрүүдийн ялгаа (дивергенц) хоорондын хугацааг харгалзан бүтээдэг. Эцэст нь, филогенетик модны график дүрслэл нь балар эртний Европын хронотоп дахь гаплогрупп I-ийн тээвэрлэгчдийн хөдөлгөөнийг коньюнктурын сэргээн босголтоор нөхөж байна. I2a хаплогруппын хуваагдлын он дараалал нь дараах байдалтай байна (бидний хувилбар): Ойролцоогоор 14,000-15,000 жилийн өмнө I2a L460/PF3647/S2381салбар бий болсон I2a2-L35/PF3862/S150, L37/PF6900/S153, L181, M436/P214/PF3856/S33, P216/PF3855/S30, P217/PF3854/S28/S231, энэ нь бараг тэр даруй хуваагдсан I2a2a-L34/PF3857/S151, L36/S152, L59, L368, L622, M223, P219/PF3859/S24, Үүнтэй зэрэгцэн I2a гаплогруппээс дэд салбар гарч ирэв I2a1-P37.2 (одоогоос 15,000-10,000 жилийн өмнө). Өргөтгөсөн шинжилгээний явцад I2a1 тусдаа гаплогруппийг тодорхойлсон нь анхаарал татаж байна -F (ISOGG модонд ороогүй L1294 L1297 L1298 зүсэгчийг тодорхойлох). Нордтведтын тооцоолсон үе хоорондын насны дагуу энэ кластер нь I2a1a бүлгээс хожуу, харин I2a1b бүлгээс эрт (өөрөөр хэлбэл, хоёр салааны дунд) үүссэн. Нордтведтийн хэлснээр I2a1-ийн сүүлчийн нийтлэг өвөг дээдэс-F ба I2a1a бидний үеэс 20 мянган жилийн өмнө амьдарч чадах байсан. I2a1*-F-ийн төлөөлөгчид ихэвчлэн орчин үеийн баруун Францын нутаг дэвсгэрт амьдардаг нь онцлог юм. Үүнээс гадна I2a1c-ийн төлөөлөгчид Баруун Европт амьдардаг. -аас I2a1-P37.2МЭӨ 20,000-15,000 оны хоорондхайрцаг түүний цаг хугацаа онцгой байвI2a1aL158/PF4073/S433, L159.1/S169.1, M26/PF4056((сүүлийн нийтлэг өвөг дээдсийн нас - - 8,000-6,000) болонI2a1b-M423 L178/S328, M423(сүүлчийн нийтлэг өвөг дээдсийн нас одоогоос 14,000-аас доошгүй жилийн өмнөх) I2a1b-M423 L178/S328хоорондын дансны дагуу өвөг дээдсийн удам угсаа 13,500 жилийн өмнө ялгагджээ.I2a1b3 ба I2a1b2 - (Dinaric+Disles) ба арлууд. Сонирхолтой нь бид арлуудын сүүлчийн нийтлэг өвөг дээдсийн насыг 6600 жилийн өмнө одоогийнхоос ойролцоогоор 8000 жил гэж тооцоолж байна.
-аас илүү хожуу үе доторх хэлтэсүүд (атяг 8 мянга-6 мянган жилийн өмнөх) ойролцоогоор дэлхийн хөргөлттэй сайн уялдаж байна. МЭӨ 6200 он бөгөөд энэ нь Европ дахь хүн ам илт буурсан үед неолитын хүн ам зүйн хямралд хүргэсэн гэж шинжээчид тэмдэглэжээ.Хойд Европт нэгэн зэрэгминий үед (ойролцоогоор МЭӨ 6500-6200 он) газарзүйн хувьд тусгаарлагдсан (филогенетикийн хувьд эрт тусгаарлагдсан) популяцууд I2a1b2-Арлууд ба I2a1b3 болсон үйл явдал болсон. , түүнчлэн үлдсэн I2a2 дэд бүлгүүдээс I2a2-M284 – Арлын дэд хэсгийн нэг хэсэг.Бид Доггерландын үерийн тухай ярьж байна. -аар тодорхойлсонСүүлийн мөстлөгийн үеэр Хойд тэнгис ба Ла-Маншийн сувгийн хоорондох усны хагалбар нь Пас-де-Калегаар биш, Зүүн Англиас зүүн өмнөд Хоек ван Холланд хүртэл урсаж байсныг нотлох баримтууд, тэр дундаа орчин үеийн далайн ёроолын тойргийг харуулсан бөгөөд Темза голууд, Меуз, Шелдт, Рейн хоёр нэгдээд Английн сувгийн дагуу өргөн гол хэлбэрээр урсаж, эцэст нь Атлантын далайд цутгаж байв.[ МЭӨ 8000 он. д. хуурай газрын хойд эрэг, Доггеrland нь далайн эрэг, далайн эрэг, намаг, далайн эрэг, далайн эрэг бүхий далайн эрэг байв. Магадгүй мезолитийн үед энэ газар ан агнуур, шувууны аж ахуй, загас агнуурын хувьд Европ дахь хамгийн баян газар байсан байж магадгүй юм. Мөсөн гол хайлсны үр дүнд далайн түвшин аажмаар нэмэгдсэн (сүүлчийн мөстлөгийн төгсгөл) нь Хойд тэнгисээр Доггерландыг усанд автуулж, балар эртний Британийг 6500 орчим Европ тивээс аажмаар таслав гэж таамаглаж байна. МЭӨ. д. Одоогийн байдлаар Доггер банк гэгддэг ирээдүйн элсэн эрэг дээр энэхүү таамаглалын дагуу арал МЭӨ 5000 он хүртэл оршин тогтнож байсан. д. Саяхан дэвшүүлсэн өөр нэг таамаглал бол 8200 жилийн өмнө (МЭӨ 6200) газар нутгийн ихэнх хэсэг нь цунамигийн улмаас усанд автсан гэсэн таамаглал юм. түүхэн дэх хамгийн томуудын нэгхөрсний гулгалт - усан доорх хөрсний талбайн эргээс шилжилт хөдөлгөөнНорвеги улсыг Стурегга гэдэг . Энэ онолын дагуу хөрсний гулгалтаас үүдэлтэй цунами нь тухайн нутгийн мезолитийн үеийн хүн амд сүйрлийн үүрэг гүйцэтгэсэн. Цунамигийн улмаас Их Британи тивээс бүрмөсөн тусгаарлагдсан мэт харагдаж байгаа бөгөөд соёлын үүднээс авч үзвэл,Мезолит өөрийнхөөрөө явсан. Гаж нөлөөний нэгхөрсний гулгалт болсондэлхийн хөргөлт МЭӨ 6200 д. хайлсан мөсөн голоос хүйтэн ус орж ирсний улмаас . I2a хаплогруппын эртний төлөөлөгчид хэн байсан бэ? I. I2a хаплогруппын өвөг дээдэс хаана гарч ирсэн бэ?? Википедиа дээрх ишлэлээс харахад зарим генетикийн хүрээний дунд I2a нь Балканы неолитын соёлын хүрээ, тэр дундаа Трипиллианчуудын ойролцоох соёлтой холбоотой гэсэн буруу үзэл баримтлалыг бэхжүүлж байна. Энэ сонголт боломжтой боловч эцсийн хувилбарыг эдгээр соёлын төлөөлөгчдийн үлдэгдлийн ДНХ-ээс хуучин Y-хромосомын шинжилгээгээр тодорхойлно. Гэхдээ хэрэв I2 нь трипиллианчуудын дунд байсан бол тэдгээрийг уугуул элемент гэж үзэх боломжгүй (тэдгээрийн цөм нь Ойрхи Дорнодоос ирсэн цагаачид байсан); тэд энэ соёлд шингэсэн орон нутгийн мезолитийн элементийн үлдэгдлийг төлөөлсөн байх магадлалтай. Хэрэв бид эртний I2a2-ийн эртний соёлын тухай ярих юм бол эдгээр нь Свидерчууд ба тэдгээрийн эпигонууд байсан байх. 2011 онд би Польшийн FTDNA төслийн администратортай (Ларри Майка) I2a1b-M423 (мөн ерөнхийдөө I2a) гаплогруппын гарал үүслийн талаар бага зэрэг ярилцаж чадсан. Үүнээс өмнө Майк энэ хаплогруппын гарал үүслийг харьцангуй бага сонирхдог байсан (хэдийгээр тэр миний нэгэн адил түүнд харьяалагддаг). Тэрээр I2a1 хаплогруппын гарал үүслийн талаар маш сонирхолтой хувилбарыг санал болгов. Түүний бодлоор өндөр магадлалтай бүх I2a1 (I2a1*, I2a1a, I2a1b, I2a1c гэх мэт) нийтлэг өвөг нь Альпийн бүсэд амьдарч болох байсан. Тэрээр мөн миний санал болгож буй I2a1 овгийг Альпийн нуруунаас Карпат руу дайран өнгөрөх замыг зөвшөөрсөн. Тэр миний үерхэхийг зөвшөөрөөгүй нь үнэн. I2a1-ийн өвөг дээдсийн хүн ам Альпийн нуруунаас Карпатын нуруу руу (баруун Европоос хойд зүг рүү ухарч буй мөсөн голын дараа) нүүдэллэж эхэлсэн гэж би итгэж байна, ЗБНХ дууссаны дараа (өөрөөр хэлбэл МЭӨ 10,000 жилийн дараа мезолитийн үед) гэж Майк үзэж байна. нүүдлийн санаачлагч нь Кельтүүд байж болох юм (өөрөөр хэлбэл энэ нүүдэл нь хүрэл зэвсгийн үед аль хэдийн болсон). Сүүлчийн сонголт нь сонирхолтой боловч арлын дэд бүлэг I2a1b2 ба Динарийн дэд бүлэг I2a1b3 тусгаарлагдсан цаг хугацаа, газрыг тайлбарлаагүй бөгөөд энэ нь молекулын болзооны дагуу 10-8 мянган жилийн өмнө тохиолдох боломжгүй юм. Майкигийн хувилбарыг шалгахын тулд би Mesquite алгоритмыг ашиглахаар шийдсэн бөгөөд энэ нь одоогийн газарзүйн тархалт (газарзүйн) дээр үндэслэн дэд бүлгийн "суурь" хамгийн их магадлалтай (парсимонийн үүднээс) газрыг тооцоолох боломжийг олгодог. тархалт) хоёр зэргэлдээ дэд бүлгүүдийн (энэ тохиолдолд бид I2a1b1 гарч ирэх газрыг гаплотип таксоны газарзүйн координат ба I2a1a гадна бүлгийн үндэсээр үндэслэсэн I2a1b модны топологийн харьцуулалт дээр үндэслэн тодорхойлно). Одоогийн байдлаар I2a1a дэд бүлгийн хамгийн өндөр нягтрал нь Сардинид байгаа хэдий ч Нордтведтын үндэслэлтэй, эмпирикээр батлагдсан дүгнэлтийн дагуу хамгийн эртний I2a1a кластерууд хойд бүс нутагт байдаг. Иберийн хойг, Альп, Францын өмнөд хэсэг. Энэ зорилгын үүднээс би Мескит дэх филогенетик модыг 2D-3D проекцоор Хойд Европын газарзүйн төсөөлөлд шууд хэрхэн ашиглах талаар сурсан. Жишээлбэл, миний Mesquite Cartographer-д бүтээсэн I2a1b-M423 687 гаплотипийн филогенетик модны бүтцэд үндэслэсэн 2 газрын зураг нь TNT дээрх 20 шилдэг (парсимоник) модны 20 бие даасан тохирохыг олж илрүүлсэн. Улаан өнгө нь сэргээн босгосон газар нутгийг илэрхийлдэг - энэ тохиолдолд Альпийн бүс.
Тиймээс, I2a1b тээвэрлэгчдийн олж авсан замнал нь бидний санал болгож буй маршруттай ойролцоо байна. I2a1b ба I2a1b3 харагдах хооронд хэдэн мянган жил өнгөрсөн тул нүүдэл яг хэзээ болсныг хэлэхэд хэцүү байдаг. Өөр нэг чухал зүйл бол хөдөлгөөн Балканы нутгаас биш, харин Франц эсвэл Швейцарийн Альпийн нуруунаас зүүн хойд зүгт эхэлсэн гэдгийг бид харуулсан. Энэ хувилбар нь эртний ДНХ гэх мэт материаллаг нотлох баримтаар батлагдсан уу? Саяхныг хүртэл бидний дэвшүүлсэн таамаглал зөвхөн статистикийн интерполяци хэвээр байсан бөгөөд нэмэлт баталгаажуулалт шаардлагатай байв. Хэдэн долоо хоногийн өмнө "мэргэжлийн" генетикчдийн бүтээл эцэст нь хэвлэгдэн гарсан нь өнөөг хүртэл судлагдсан эртний дээжүүдийн ихэнх ДНХ нь орчин үеийн хүний популяцийн ДНХ-тэй хэрхэн уялдаж байгааг харуулж байна ( Эртний хүний геномууд нь одоогийн Европчуудын гурван өвөг дээдсийн популяцийг санал болгодог). Өнгөрсөн жилүүдэд миний хэлсэн I2a1b-ийн нүүдлийн эхлэл, эхлэлийн талаархи миний таамаглал энэ ажилд материаллаг нотлогдож байна. Нэгдүгээрт, ганц эцэг эхийн шугамаар Швед, Люксембургийн мезолитийн анчин цуглуулагчдын эртний ДНХ-ийн генийн сан нь mtDNA гаплогрупп U ба Y-хромосомын гаплогрупп I-ийн төлөөлөл болж хувирсан тул бид дахин эрт үед R1a/R1b байхгүйтэй тулгарч байна. Мезолитийн Европчууд. Хоёрдугаарт, эдгээр судалгаанууд үүнийг харуулж байна ЮксембургМезолитийн анчин цуглуулагч (түүний шарилыг одоогоос 8000 жилийн өмнө Люксембургийн Хеффинген-Лохбургийн мезолитийн үеийн хайгуулын үеэр олсон) хаплогруппын бүлэгт багтдаг байв. I2a1b* M423 + L178+. Хүснэгтэд өгөгдсөн Y-хромосомын полиморфизмын талаарх мэдээллээс үзэхэд энэхүү эртний Люксембургийн гаплогрупп нь I2a1b2 ба I2a1b3-тай холбоотой өвөг дээдэс юм. I2a1b L178 G A 15574052 12 + Гуравдугаарт, эдгээр судалгаанаас үзэхэд Моталагийн мезолитийн үеийн хүн амын нэг хэсэг (Швед, Өстгетерланд, одоогоос 8000 орчим жилийн өмнө) мөн адил харъяалагдаж байсныг харуулж байна. I2a1b*(xI2a1b1, I2a1b3) |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||






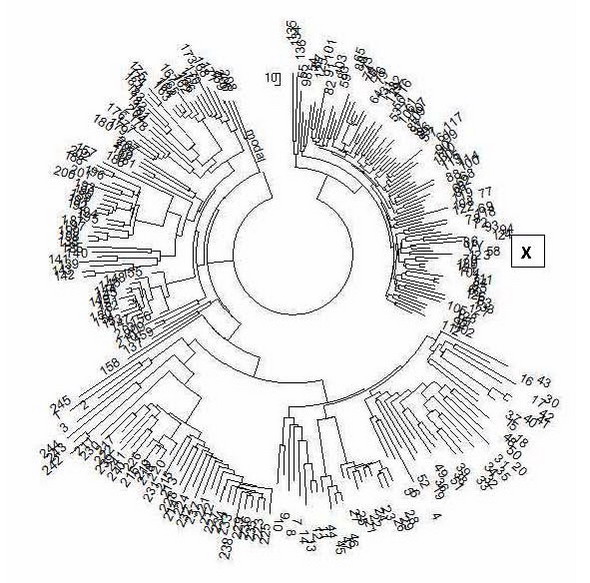













Алдартай:

Шинэ
- Ял шийтгүүлсэн хүнийг нийгэмшүүлэх 14 үндсэн чиглэл
- Вячеслав Наговицын ажил олдсонгүй
- Алексей Пушков: намтар, хувийн амьдрал, гэр бүл, гэрэл зураг Хүүхэд нас, гэр бүл
- Печерскийн Гэгээн Сисой, схем лам Печерскийн Ариун Эрхэм лам Иосаф
- Есүс Христийн амилалт Христийн амилалтын дараа юу болсон
- Ариун онгон Мариагийн сүм Ариун онгон Мариагийн сүм
- Ортодокс нэвтэрхий толь бичгийн мод дахь Дөрөв дэх Менионы утга
- Эрүүл мэндийн яамны 02-ны өдрийн 290н тоот тушаал
- Экспоненциал тэгш бус байдал
- Тогтсон стандартыг үндэслэн эмч нарын ажлын ачааллын стандарт, орон тооны стандарт болон бусад хөдөлмөрийн стандартыг тооцдог.




